

Reprogramming cellular events by poly(ADP-ribose)-binding proteins
- PMID: 23268355
- PMCID: PMC3812366
- DOI: 10.1016/j.mam.2012.12.005
Reprogramming cellular events by poly(ADP-ribose)-binding proteins
Abstract
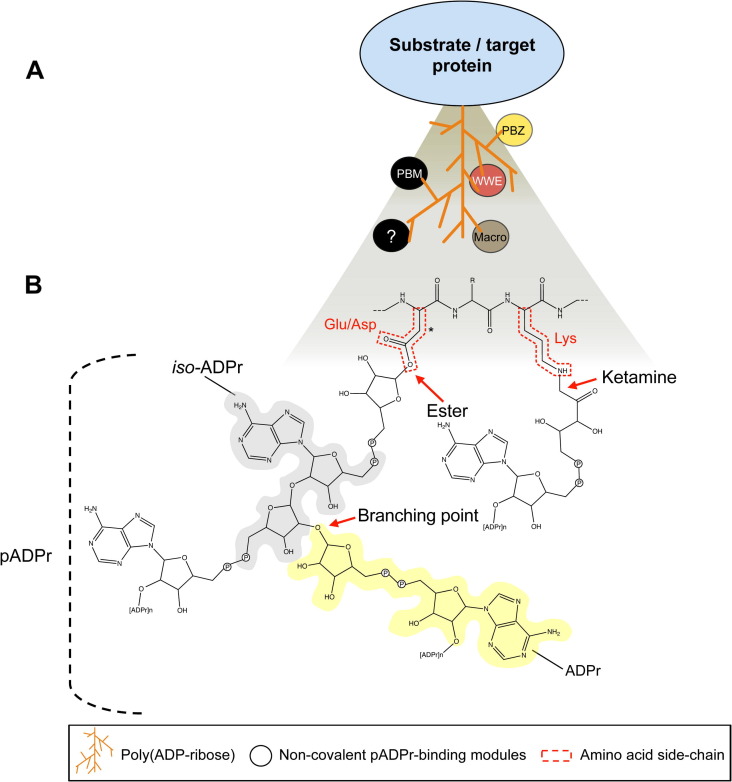
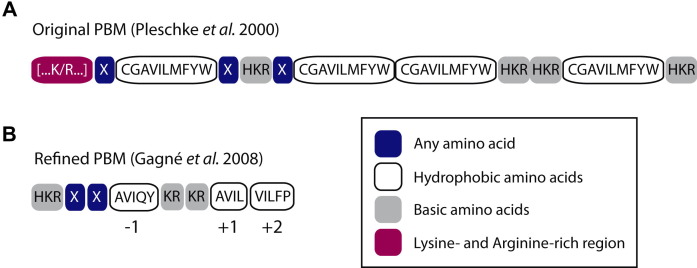
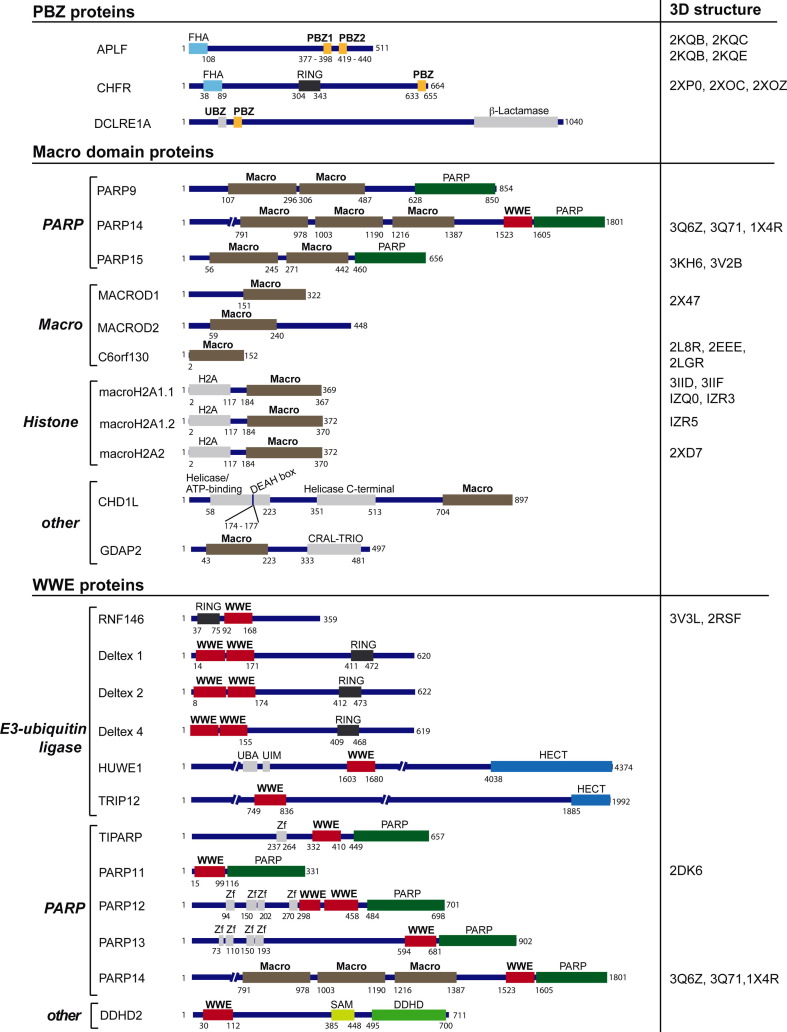
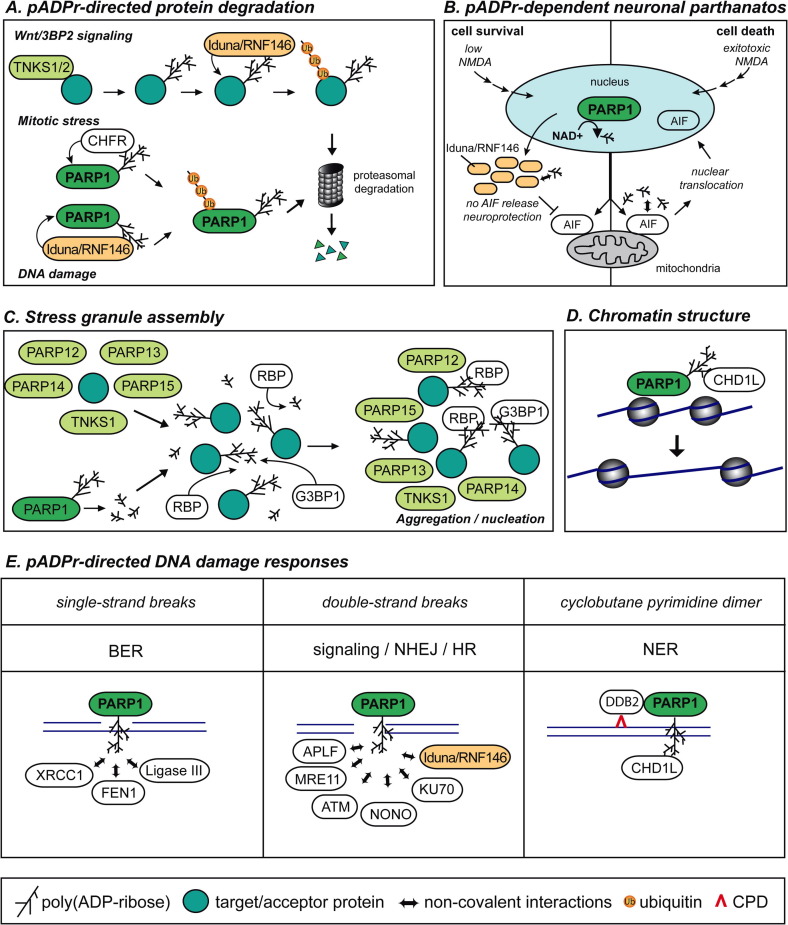
Poly(ADP-ribosyl)ation is a posttranslational modification catalyzed by the poly(ADP-ribose) polymerases (PARPs). These enzymes covalently modify glutamic, aspartic and lysine amino acid side chains of acceptor proteins by the sequential addition of ADP-ribose (ADPr) units. The poly(ADP-ribose) (pADPr) polymers formed alter the physico-chemical characteristics of the substrate with functional consequences on its biological activities. Recently, non-covalent binding to pADPr has emerged as a key mechanism to modulate and coordinate several intracellular pathways including the DNA damage response, protein stability and cell death. In this review, we describe the basis of non-covalent binding to pADPr that has led to the emerging concept of pADPr-responsive signaling pathways. This review emphasizes the structural elements and the modular strategies developed by pADPr-binding proteins to exert a fine-tuned control of a variety of pathways. Poly(ADP-ribosyl)ation reactions are highly regulated processes, both spatially and temporally, for which at least four specialized pADPr-binding modules accommodate different pADPr structures and reprogram protein functions. In this review, we highlight the role of well-characterized and newly discovered pADPr-binding modules in a diverse set of physiological functions.
Keywords: Macro domain; PARG; PARP; PBZ; Poly(ADP-ribose); WWE.
Copyright © 2013 Elsevier Ltd. All rights reserved.
Figures
References
-
- Adamietz P., Oldekop M., Hilz H. Proceedings: is poly(ADP-ribose) covalently bound to nuclear proteins? J. Biochem. 1975;77:4p. - PubMed
-
- Adhikary S., Marinoni F., Hock A., Hulleman E., Popov N., Beier R., Bernard S., Quarto M., Capra M., Goettig S., Kogel U., Scheffner M., Helin K., Eilers M. The ubiquitin ligase HectH9 regulates transcriptional activation by Myc and is essential for tumor cell proliferation. Cell. 2005;123:409–421. - PubMed
-
- Aguiar R.C., Takeyama K., He C., Kreinbrink K., Shipp M.A. B-aggressive lymphoma family proteins have unique domains that modulate transcription and exhibit poly(ADP-ribose) polymerase activity. J. Biol. Chem. 2005;280:33756–33765. - PubMed
-
- Aguiar R.C., Yakushijin Y., Kharbanda S., Salgia R., Fletcher J.A., Shipp M.A. BAL is a novel risk-related gene in diffuse large B-cell lymphomas that enhances cellular migration. Blood. 2000;96:4328–4334. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
