

doi: 10.1128/JVI.01292-12.
Epub 2012 Dec 26.
Unraveling of a neutralization mechanism by two human antibodies against conserved epitopes in the globular head of H5 hemagglutinin
Affiliations
- PMID: 23269809
- PMCID: PMC3592130
- DOI: 10.1128/JVI.01292-12
Item in Clipboard
Unraveling of a neutralization mechanism by two human antibodies against conserved epitopes in the globular head of H5 hemagglutinin
J Virol.
2013 Mar.
Abstract
The rapid spread of highly pathogenic avian influenza (HPAI) H5N1 virus underscores the importance of effective antiviral treatment. Previously, we developed human monoclonal antibodies 65C6 and 100F4 that neutralize almost all (sub)clades of HPAI H5N1. The conserved 65C6 epitope was mapped to the globular head of HA. However, neither the 100F4 epitope nor the neutralization mechanism by these antibodies was known. In this study, we determined the 100F4 epitope and unraveled a neutralization mechanism by antibodies 65C6 and 100F4.
Figures
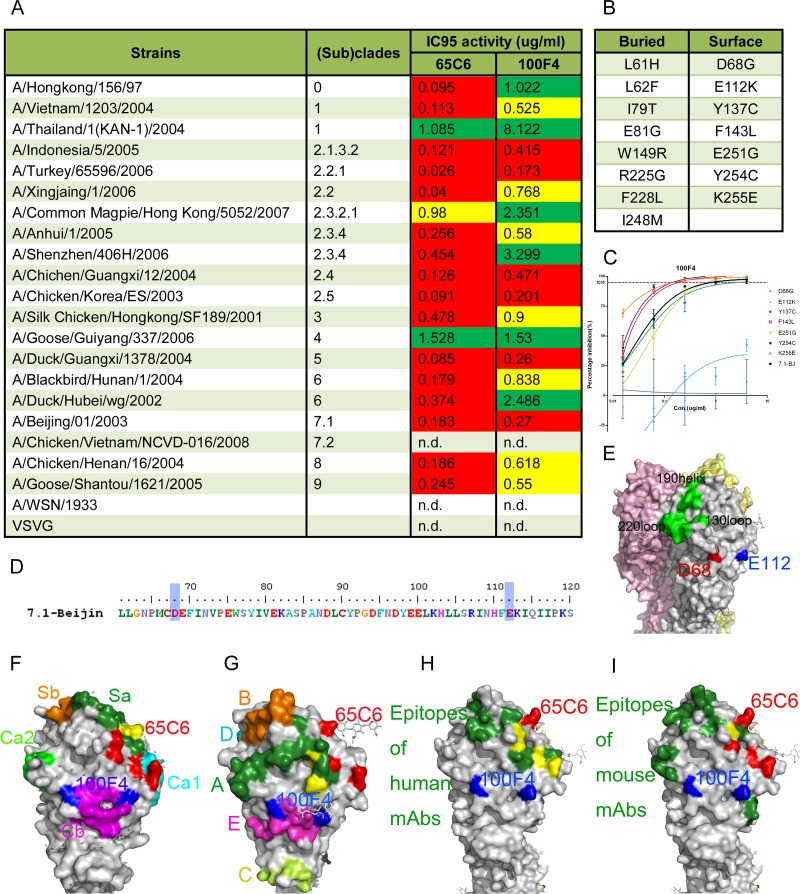
Amino acid residues involved in the neutralization epitope of 100F4. (A) Broad neutralization activity (95% inhibitory concentration [IC95]) of antibodies 65C6 and 100F4 against a panel of H5N1 pseudotypes reproduced from reference . Green, >1 μg/ml required to reach IC95; yellow, between 0.5 and 1 μg/ml required to reach IC95; red, <0.5 μg/ml required to reach IC95. (B) List of 15 single amino acid mutants that antibody 100F4 can no longer bind to, obtained by using yeast Saccharomyces cerevisiae that displays a random mutagenesis library of HA fragment comprising amino acid residues 51 to 260 in fine epitope mapping. Among them, 8 amino acid mutants are underneath the HA surface and the other 7 are on the surface of HA. (C) Titration of antibody 100F4 against H5N1 pseudotypes expressing 7 single surface amino acid mutants compared to the results against H5N1 pseudotype expressing the parental HA. (D) Amino acid residues D68 and E112 (D72 and E116 in H3 numbering) involved in 100F4 epitope are highlighted by blue shading. (E) Amino acid residues D68 and E112 are highlighted in red and blue, respectively, in space-filling model of HA. Yellow, pink, and grey each indicate one of three monomers that make up an HA trimer. (F) 100F4 and 65C6 epitopes in the context of known neutralization epitopes in the H1 HA structure (Protein Data Bank [PDB] structure accession number IRU7), as follows: Ca1, cyan; Ca2, light green; Cb, magenta; Sa, forest green; and Sb, orange. Yellow, overlap amino acid residues between the 65C6 epitope and the Sa site. (G) 100F4 and 65C6 epitopes in the context of known neutralization epitopes in H3 HA structure (PBD structure accession number 2VIU), as follows: site A, forest green; site B, orange; site C, yellow-green; site D, cyan; and site E, magenta. Yellow, overlap amino acid residues between the 65C6 epitope and site A. (H) 100F4 and 65C6 epitopes in the context of known neutralization epitopes detected by human MAb in H5 HA structure (PBD structure accession number 2ibx). Neutralization epitopes detected by human MAb are highlighted in forest green, and amino acid residues that overlap amino acid residues of the 65C6 epitope are highlighted in yellow. (I) 100F4 and 65C6 epitopes in the context of known neutralization epitopes detected by mouse MAb in H5 HA structure (PBD structure accession number 2ibx). Neutralization epitopes detected by mouse MAb are highlighted in forest green, and amino acid residues that overlap amino acid residues of 65C6 epitope are highlighted in yellow.
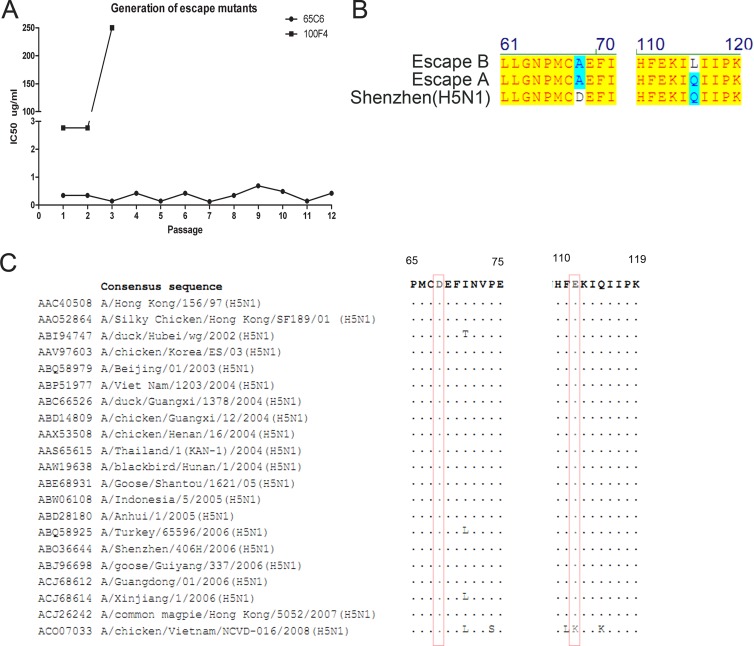
Determination of antibody-driven escape mutants. (A) Antibody-driven escape mutants were generated through a series of passages. (B) Point mutations found in two 100F4 antibody-driven escape clones compared to the sequence of parental H5 HA. (C) Sequence alignment of amino acid residues 65 to 75 and 110 to 119 among 21 HA covering all H5 clades and subclades. Residues 68 and 112 are highlighted in boxes.
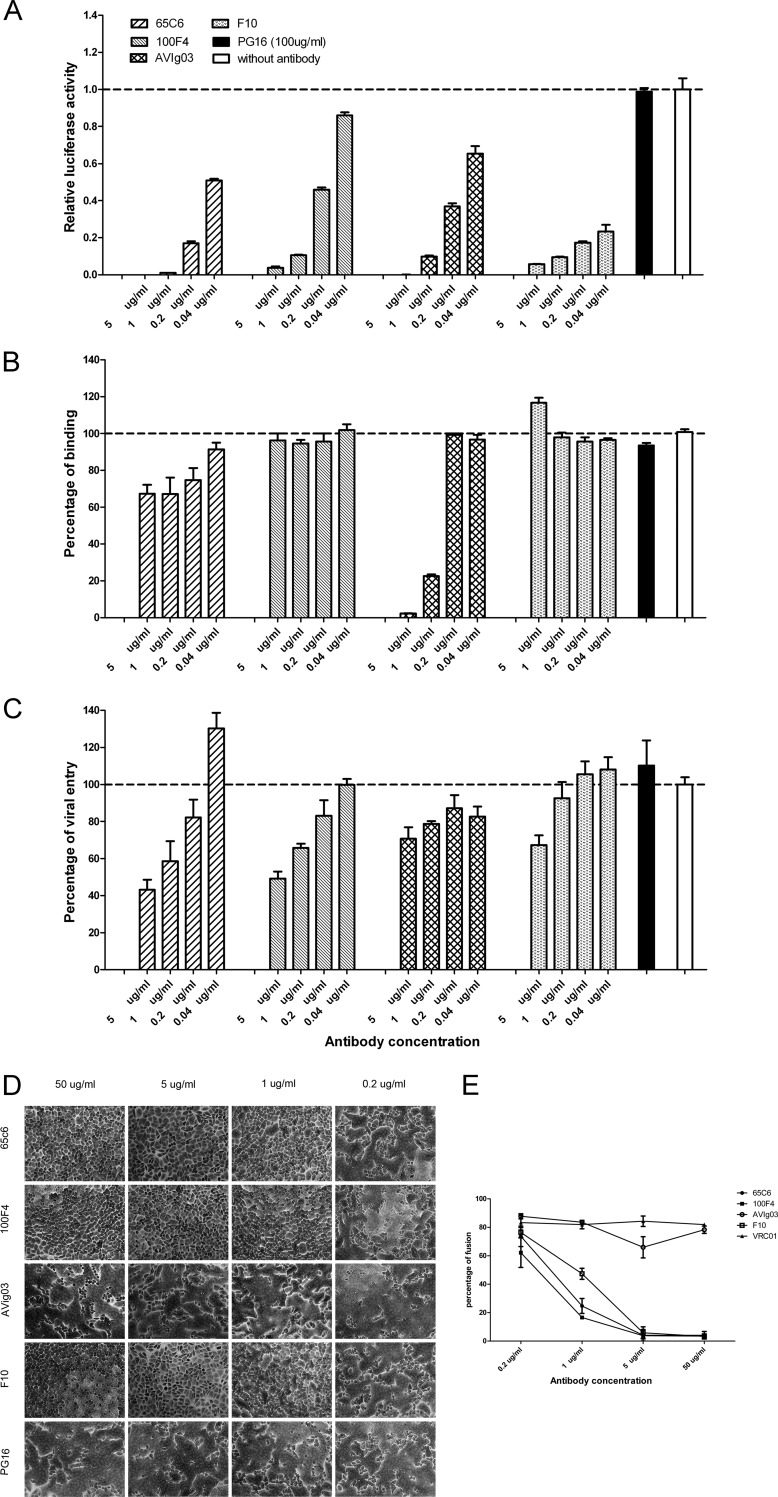
Determination of effects of antibodies 65C6 and 100F4 on virus neutralization, attachment, postattachment, and fusion. (A) Neutralization activity against H5N1 pseudotype A/Shenzhen/406H/06 by indicated concentrations of antibodies 65C6, 100F4, AVIg03, F10, and PG16. The neutralization activity was calculated as follows: relative luciferase activity (RLA) of pseudotype with a given concentration of an antibody/RLA of pseudotype alone without antibodies. (B) Effects of indicated concentrations of antibodies 65C6, 100F4, AVIg03, F10, and PG16 on binding of H5N1 pseudotype A/Shenzhen/406H/06 to MDCK cells compared to binding of pseudotype alone (without antibodies). (C) Effects of indicated concentrations of antibodies 65C6, 100F4, AVIg03, F10, and PG16 on postattachment entry of H5N1 pseudotype A/Shenzhen/406H/06 into MDCK cells compared to entry of pseudotype alone (without antibodies). (D) Effects of antibodies 65C6, 100F4, AVIg03, F10, and PG16 on pH 5.0-triggered, HA-mediated cell-cell fusion among influenza H5 HA-transduced HeLa cells. (E) Mean and standard deviation of fusion percentages in each image in panel D. The fusion percentages were calculated as described in the text.
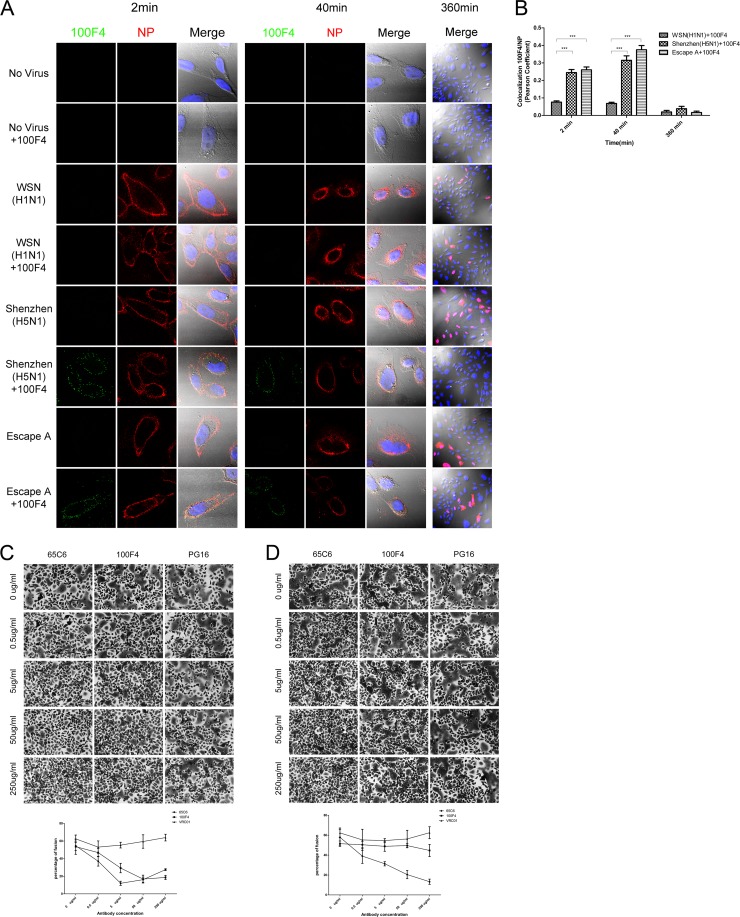
Bound antibodies 65C6 and 100F4 are internalized and block low-pH-induced fusion. (A) Confocal microscopy of virus internalization into CHO-K1 cells with or without antibody 100F4. Cell samples were collected at 2, 40, and 360 min postinfection as indicated. Green, antibody 100F4 staining; red, viral NP staining; blue, DAPI staining for nucleus. (B) Colocalization of antibody 100F4 and viral NP, as determined by Pearson's correlation coefficients, was quantified at 2, 40, and 360 min postinfection. ***, P < 0.001. (C) Blockage of low-pH-triggered, wild-type H5 HA-mediated fusion by antibodies 65C6 and 100F4 but not PG16 control. Mean and standard deviation of fusion percentage in each image is shown below. (D) Blockage of low-pH-triggered, escape A of H5 HA-mediated fusion by antibody 65C6 but not by antibody 100F4 and PG16 control. Mean and standard deviation of fusion percentage in each image is shown below.
Similar articles
-
Divergent Requirement of Fc-Fcγ Receptor Interactions for In Vivo Protection against Influenza Viruses by Two Pan-H5 Hemagglutinin Antibodies.J Virol. 2017 May 12;91(11):e02065-16. doi: 10.1128/JVI.02065-16. Print 2017 Jun 1. J Virol. 2017. PMID: 28331095 Free PMC article.
-
A human antibody recognizing a conserved epitope of H5 hemagglutinin broadly neutralizes highly pathogenic avian influenza H5N1 viruses.J Virol. 2012 Mar;86(6):2978-89. doi: 10.1128/JVI.06665-11. Epub 2012 Jan 11. J Virol. 2012. PMID: 22238297 Free PMC article.
-
An antibody against a novel and conserved epitope in the hemagglutinin 1 subunit neutralizes numerous H5N1 influenza viruses.J Virol. 2010 Aug;84(16):8275-86. doi: 10.1128/JVI.02593-09. Epub 2010 Jun 2. J Virol. 2010. PMID: 20519402 Free PMC article.
-
Recent advances in "universal" influenza virus antibodies: the rise of a hidden trimeric interface in hemagglutinin globular head.Front Med. 2020 Apr;14(2):149-159. doi: 10.1007/s11684-020-0764-y. Epub 2020 Apr 1. Front Med. 2020. PMID: 32239416 Free PMC article. Review.
-
The antigenic architecture of the hemagglutinin of influenza H5N1 viruses.Mol Immunol. 2013 Dec;56(4):705-19. doi: 10.1016/j.molimm.2013.07.010. Epub 2013 Aug 7. Mol Immunol. 2013. PMID: 23933511 Review.
Cited by
-
Structural and functional definition of a vulnerable site on the hemagglutinin of highly pathogenic avian influenza A virus H5N1.J Biol Chem. 2019 Mar 22;294(12):4290-4303. doi: 10.1074/jbc.RA118.007008. Epub 2019 Feb 8. J Biol Chem. 2019. PMID: 30737282 Free PMC article.
-
Complementary recognition of the receptor-binding site of highly pathogenic H5N1 influenza viruses by two human neutralizing antibodies.J Biol Chem. 2018 Oct 19;293(42):16503-16517. doi: 10.1074/jbc.RA118.004604. Epub 2018 Aug 28. J Biol Chem. 2018. PMID: 30154240 Free PMC article.
-
B Cell Responses against Influenza Viruses: Short-Lived Humoral Immunity against a Life-Long Threat.Viruses. 2021 May 22;13(6):965. doi: 10.3390/v13060965. Viruses. 2021. PMID: 34067435 Free PMC article. Review.
-
Crimean-Congo haemorrhagic fever virus.Nat Rev Microbiol. 2023 Jul;21(7):463-477. doi: 10.1038/s41579-023-00871-9. Epub 2023 Mar 14. Nat Rev Microbiol. 2023. PMID: 36918725 Free PMC article. Review.
-
Association of poultry vaccination with interspecies transmission and molecular evolution of H5 subtype avian influenza virus.Sci Adv. 2025 Jan 24;11(4):eado9140. doi: 10.1126/sciadv.ado9140. Epub 2025 Jan 22. Sci Adv. 2025. PMID: 39841843 Free PMC article.
References
-
- Hu H, Voss J, Zhang G, Buchy P, Zuo T, Wang L, Wang F, Zhou F, Wang G, Tsai C, Calder L, Gamblin SJ, Zhang L, Deubel V, Zhou B, Skehel JJ, Zhou P. 2012. A human antibody recognizing a conserved epitope of H5 hemagglutinin broadly neutralizes highly pathogenic avian influenza H5N1 viruses. J. Virol. 86:2978–2989 - PMC - PubMed
-
- Tsai C, Caillet C, Hu H, Zhou F, Ding H, Zhang G, Zhou B, Wang S, Lu S, Buchy P, Deubel V, Vogel FR, Zhou P. 2009. Measurement of neutralizing antibody responses against H5N1 clades in immunized mice and ferrets using pseudotypes expressing influenza hemagglutinin and neuraminidase. Vaccine 27:6777–6790 - PMC - PubMed
-
- Caton AJ, Brownlee GG, Yewdell JW, Gerhard W. 1982. The antigenic structure of the influenza virus A/PR/8/34 hemagglutinin (H1 subtype). Cell 31:417–427 - PubMed
-
- Stray SJ, Pittman LB. 2012. Subtype- and antigenic site-specific differences in biophysical influences on evolution of influenza virus hemagglutinin. Virol. J. 9:91 doi:10.1186/1743-422X-9-91 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
