

Review
doi: 10.1098/rsob.120140.
Exotic mitotic mechanisms
Affiliations
- PMID: 23271831
- PMCID: PMC3603444
- DOI: 10.1098/rsob.120140
Item in Clipboard
Review
Exotic mitotic mechanisms
Open Biol.
.
Abstract
The emergence of eukaryotes around two billion years ago provided new challenges for the chromosome segregation machineries: the physical separation of multiple large and linear chromosomes from the microtubule-organizing centres by the nuclear envelope. In this review, we set out the diverse solutions that eukaryotic cells use to solve this problem, and show how stepping away from 'mainstream' mitosis can teach us much about the mechanisms and mechanics that can drive chromosome segregation. We discuss the evidence for a close functional and physical relationship between membranes, nuclear pores and kinetochores in generating the forces necessary for chromosome segregation during mitosis.
Figures
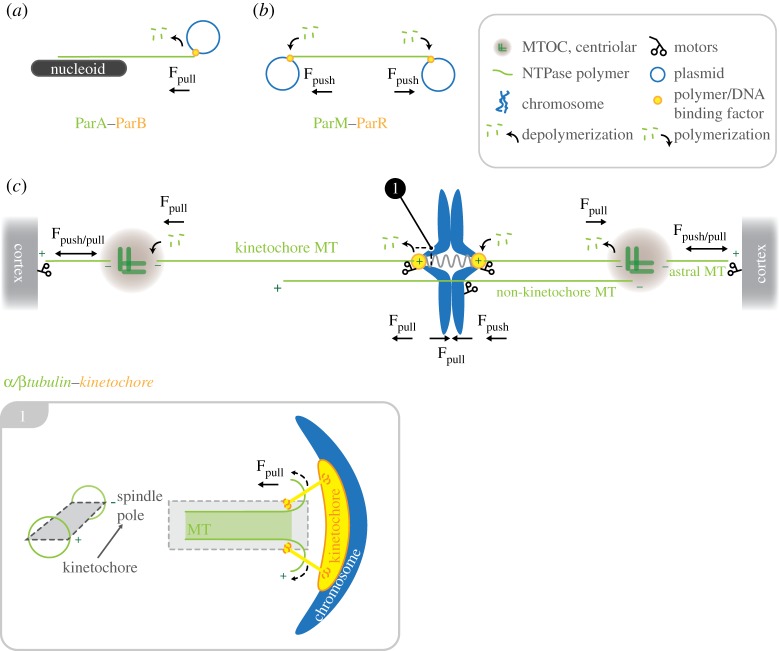
Schematic diagram showing the set-up of the chromosome segregation machinery in bacteria and mammalian cells. The basic requirement for chromosome segregation is a dynamic NTPase polymer (ParA, ParM in prokaryotes or tubulin in eukaryotes; green) that can create pushing (Fpush in a,c) or pulling forces (Fpull in b,c) by polymerization or depolymerization, respectively. Dynamic NTPase polymers are connected to DNA by centromere binding factor: ParB, ParR in prokaryotes (a,b) or the kinetochore complex in mammalian cells (c) (orange). In contrast to the simplified prokaryote machinery (a,b), chromosome segregation in eukaryotes is driven by a sum of forces created by tubulin (de)polymerization at kinetochores (plus-end) and depolymerization at spindle poles (minus-ends), in addition to MT-dependent motors bound at kinetochores, chromosome arms, microtubule-organizing centres and the cell cortex. Inset: longitudinal section through a kinetochore-bound microtubule. Curling of kinetochore-MT protofilaments at the plus-end produces pulling forces on the attached kinetochore-chromosome.

Diverse metaphase spindle set-ups in eukaryotes: (a) animals, (b) Plasmodiophora brassicae, (c) Saccharomyces cerevisiae, (d) Trypanosoma bruceri, (e) Crypthecodinium cohnii and (f) Oodinium, Amphidinium carterae. For clarity, the outline of the cell has been omitted. Free microtubule-organizing centres in (d) and (e) correspond to basal bodies that are not involved in the set-up of the mitotic spindle. Numbered insets on the right-hand side are magnifications of the MT–kinetochore–chromosome interaction sites in the corresponding overviews on the left.

Hypothetical overview of interactions and mechanisms that might mediate the insertion of membrane-bound kinetochores into the nuclear envelope (NE). (a) Summary of the mechanisms that lead to de novo insertion of nuclear pore complexes (and spindle pole bodies) into the NE. Collaborative action of transmembrane pore membrane proteins (POMs) (red) and reticulons (green) results in the convergence of the inner and outer nuclear membrane, leading to the eventual fusion of membranes. The result is a pore in the NE. Once the pore has established POMs and nucleoporins (NUPs) containing ALPS-motifs (see main text) line the highly curved membrane within the nuclear pore and provide a membrane-scaffold for further members of the nuclear pore complex (NPC) to assemble on. (b) The Nup107/160 complex is a central hub that links to the kinetochore and might mediate membrane insertion of kinetochores. Nup107/160 interacts with CENP-E and the Ndc80-complex at the kinetochore, as well as with the gamma tubulin ring complex (γ-TuRC)—a ring-like matrix that contributes to MT-nucleation. Nup160 was shown to bind to POM121 within the membranes around the nuclear pore. Nup133 was shown to contain a functional ALPS-motif (see text) that specifically binds to membranes of high curvature and is thus able to directly interact with the membrane environment within the pore. (c) Diagram of a prophase arrangement in (some) dinoflagellates in which kinetochores are inserted in the NE. The MTOC(s) have invaginated into the nucleus. Chromosomes cluster by this invagination, which is characterized by a high density of nuclear pore complexes and membranes with high curvatures. (d) Kinetochore maturing and membrane insertion in Trychonympha agilis as described by Kubai [93]. Stage I: fibrous kinetochore material, associated with a NPC, starts to assemble between the chromosome and the NE. Stage II: the kinetochore matures as a dense disc positioned between chromosome and the fibrous material. Stage III: an intermediate structure within the insertion process, resembling an intra-membrane vesicle. The dense kinetochore disc has already been inserted into the NE. The vesicle structure is filled with the filamentous material and contains at least one NPC. Stage IV: the vesicle structure has resolved. The fibrous kinetochore material is now on the cytoplasmic surface of the NE and able to accept spindle MTs.
References
-
- Gerdes K, Howard M, Szardenings F. 2010. Pushing and pulling in prokaryotic DNA segregation. Cell 141, 927–94210.1016/j.cell.2010.05.033 (doi:10.1016/j.cell.2010.05.033) - DOI - DOI - PubMed
-
- Schumacher MA. 2012. Bacterial plasmid partition machinery: a minimalist approach to survival. Curr. Opin. Struct. Biol. 22, 72–7910.1016/j.sbi.2011.11.001 (doi:10.1016/j.sbi.2011.11.001) - DOI - DOI - PMC - PubMed
-
- Salje J, Gayathri P, Löwe J. 2010. The ParMRC system: molecular mechanisms of plasmid segregation by actin-like filaments. Nat. Rev. Microbiol. 8, 683–69210.1038/nrmicro2425 (doi:10.1038/nrmicro2425) - DOI - DOI - PubMed
-
- Aylett CHS, et al. 2010. Filament structure of bacterial tubulin homologue TubZ. Proc. Natl Acad. Sci. USA. 107, 19 766–19 77110.1073/pnas.1010176107 (doi:10.1073/pnas.1010176107) - DOI - DOI - PMC - PubMed
-
- Jensen MR, Løbner-Olesen A, Rasmussen KV. 1990. Escherichia coli minichromosomes: random segregation and absence of copy number control. J. Mol. Biol. 215, 257–26510.1016/S0022-2836(05)80344-4 (doi:10.1016/S0022-2836(05)80344-4) - DOI - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources