

Evidence for small RNAs homologous to effector-encoding genes and transposable elements in the oomycete Phytophthora infestans
- PMID: 23272103
- PMCID: PMC3522703
- DOI: 10.1371/journal.pone.0051399
Evidence for small RNAs homologous to effector-encoding genes and transposable elements in the oomycete Phytophthora infestans
Abstract
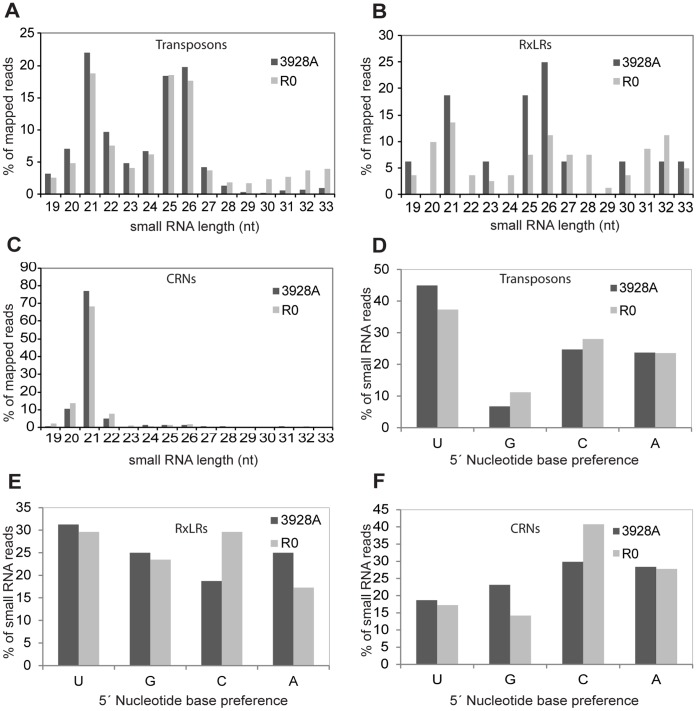
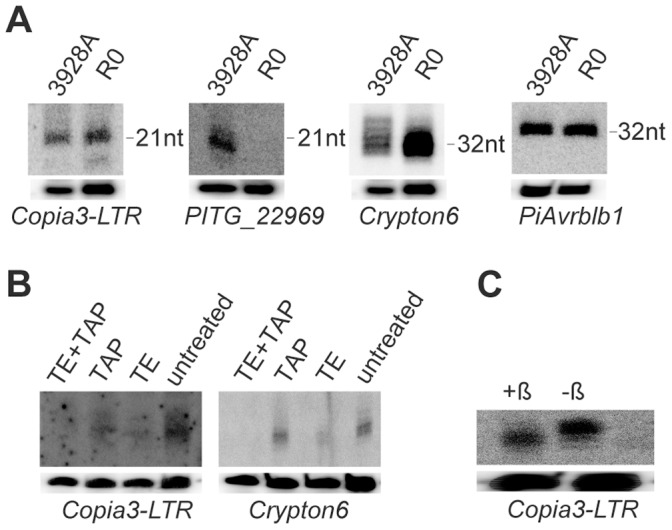
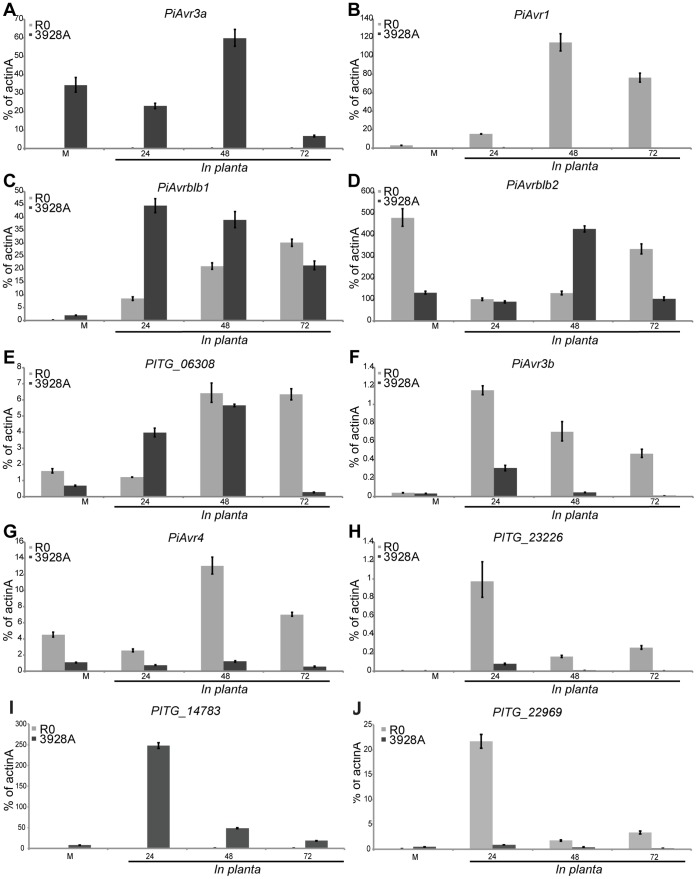
Phytophthora infestans is the oomycete pathogen responsible for the devastating late blight disease on potato and tomato. There is presently an intense research focus on the role(s) of effectors in promoting late blight disease development. However, little is known about how they are regulated, or how diversity in their expression may be generated among different isolates. Here we present data from investigation of RNA silencing processes, characterized by non-coding small RNA molecules (sRNA) of 19-40 nt. From deep sequencing of sRNAs we have identified sRNAs matching numerous RxLR and Crinkler (CRN) effector protein genes in two isolates differing in pathogenicity. Effector gene-derived sRNAs were present in both isolates, but exhibited marked differences in abundance, especially for CRN effectors. Small RNAs in P. infestans grouped into three clear size classes of 21, 25/26 and 32 nt. Small RNAs from all size classes mapped to RxLR effector genes, but notably 21 nt sRNAs were the predominant size class mapping to CRN effector genes. Some effector genes, such as PiAvr3a, to which sRNAs were found, also exhibited differences in transcript accumulation between the two isolates. The P. infestans genome is rich in transposable elements, and the majority of sRNAs of all size classes mapped to these sequences, predominantly to long terminal repeat (LTR) retrotransposons. RNA silencing of Dicer and Argonaute genes provided evidence that generation of 21 nt sRNAs is Dicer-dependent, while accumulation of longer sRNAs was impacted by silencing of Argonaute genes. Additionally, we identified six microRNA (miRNA) candidates from our sequencing data, their precursor sequences from the genome sequence, and target mRNAs. These miRNA candidates have features characteristic of both plant and metazoan miRNAs.
Conflict of interest statement
Figures



References
-
- Dick MW (2001) The Peronosporomycetes. In: McLaughlin DJ, McLaughlin EG, Lemke PA, eds. The Mycota. Vol VII. Systematics and evolution, Part A. Berlin Heidelberg New York, Springer. 39–72.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
