

Differential requirement for protein synthesis in presynaptic unmuting and muting in hippocampal glutamate terminals
- PMID: 23272190
- PMCID: PMC3521764
- DOI: 10.1371/journal.pone.0051930
Differential requirement for protein synthesis in presynaptic unmuting and muting in hippocampal glutamate terminals
Abstract
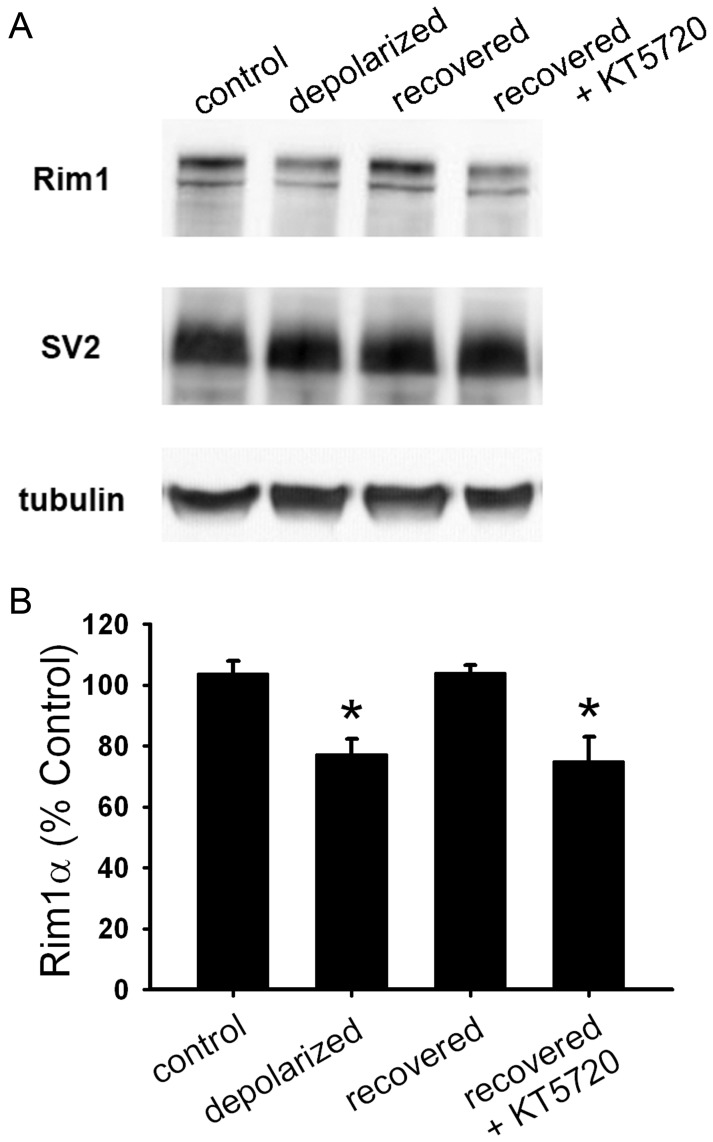
Synaptic function and plasticity are crucial for information processing within the nervous system. In glutamatergic hippocampal neurons, presynaptic function is silenced, or muted, after strong or prolonged depolarization. This muting is neuroprotective, but the underlying mechanisms responsible for muting and its reversal, unmuting, remain to be clarified. Using cultured rat hippocampal neurons, we found that muting induction did not require protein synthesis; however, slow forms of unmuting that depend on protein kinase A (PKA), including reversal of depolarization-induced muting and forskolin-induced unmuting of basally mute synapses, required protein synthesis. In contrast, fast unmuting of basally mute synapses by phorbol esters was protein synthesis-independent. Further studies of recovery from depolarization-induced muting revealed that protein levels of Rim1 and Munc13-1, which mediate vesicle priming, correlated with the functional status of presynaptic terminals. Additionally, this form of unmuting was prevented by both transcription and translation inhibitors, so proteins are likely synthesized de novo after removal of depolarization. Phosphorylated cyclic adenosine monophosphate response element-binding protein (pCREB), a nuclear transcription factor, was elevated after recovery from depolarization-induced muting, consistent with a model in which PKA-dependent mechanisms, possibly including pCREB-activated transcription, mediate slow unmuting. In summary, we found that protein synthesis was required for slower, PKA-dependent unmuting of presynaptic terminals, but it was not required for muting or a fast form of unmuting. These results clarify some of the molecular mechanisms responsible for synaptic plasticity in hippocampal neurons and emphasize the multiple mechanisms by which presynaptic function is modulated.
Conflict of interest statement
Figures






References
-
- Ninan I, Arancio O (2004) Presynaptic CaMKII is necessary for synaptic plasticity in cultured hippocampal neurons. Neuron 42: 129–141. - PubMed
-
- Moulder KL, Meeks JP, Shute AA, Hamilton CK, de Erausquin G, et al. (2004) Plastic elimination of functional glutamate release sites by depolarization. Neuron 42: 423–435. - PubMed
-
- Kohara K, Ogura A, Akagawa K, Yamaguchi K (2001) Increase in number of functional release sites by cyclic AMP-dependent protein kinase in cultured neurons isolated from hippocampal dentate gyrus. Neurosci Res 41: 79–88. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
