

A "mechanistic" explanation of the multiple helical forms adopted by bacterial flagellar filaments
- PMID: 23274110
- PMCID: PMC3605589
- DOI: 10.1016/j.jmb.2012.12.007
A "mechanistic" explanation of the multiple helical forms adopted by bacterial flagellar filaments
Abstract
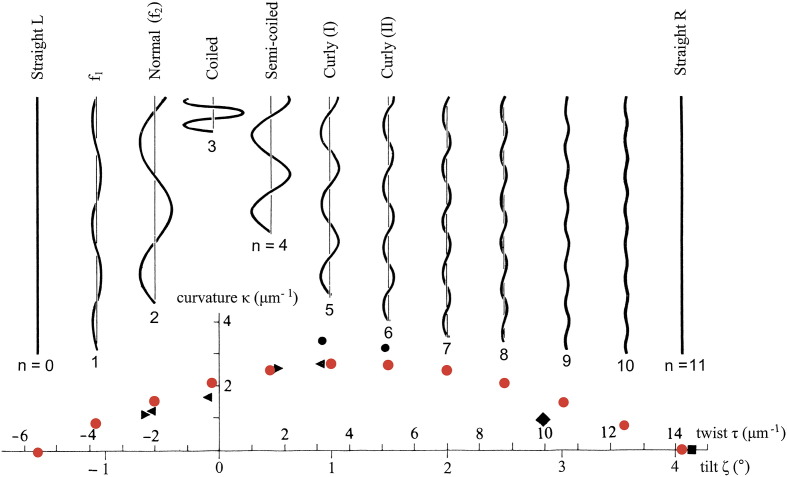
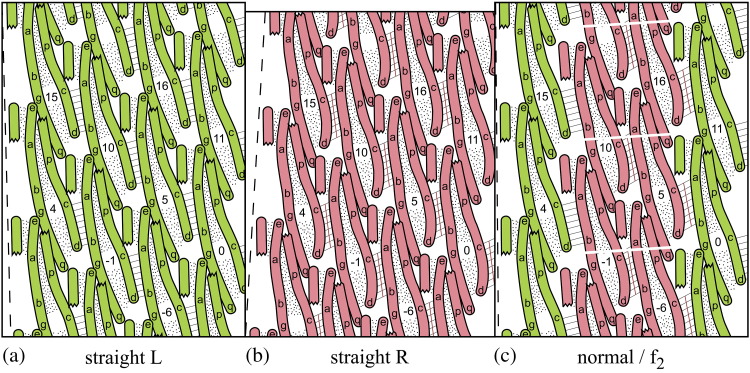
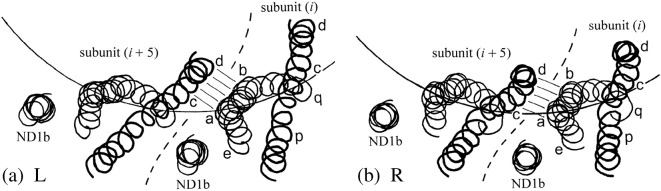
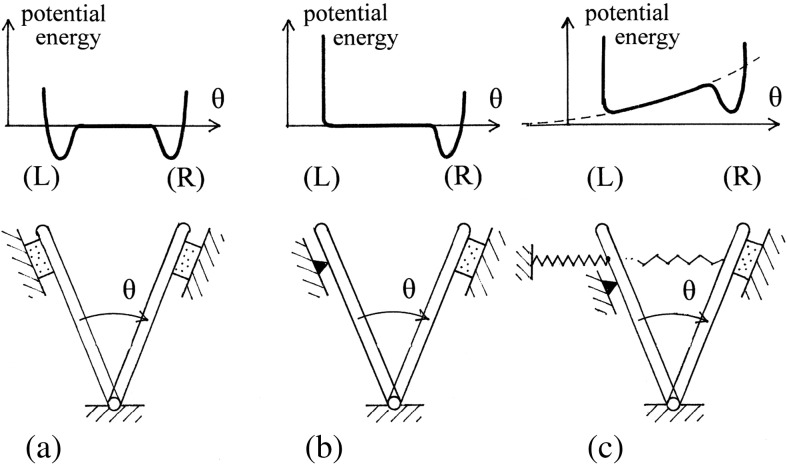
The corkscrew-like flagellar filaments emerging from the surface of bacteria such as Salmonella typhimurium propel the cells toward nutrient and away from repellents. This kind of motility depends upon the ability of the flagellar filaments to adopt a range of distinct helical forms. A filament is typically constructed from ~30,000 identical flagellin molecules, which self-assemble into a tubular structure containing 11 near-longitudinal protofilaments. A "mechanical" model, in which the flagellin building block has the capacity to switch between two principal interfacial states, predicts that the filament can assemble into a "canonical" family of 12 distinct helical forms, each having unique curvature and twist: these include two "extreme" straight forms having left- and right-handed twists, respectively, and 10 intermediate helical forms. Measured shapes of the filaments correspond well with predictions of the model. This report is concerned with two unanswered questions. First, what properties of the flagellin determine which of the 12 discrete forms is preferred? Second, how does the interfacial "switch" work, at a molecular level? Our proposed solution of these problems is based mainly on a detailed examination of differences between the available electron cryo-microscopy structures of the straight L and R filaments, respectively.
Copyright © 2012 Elsevier Ltd. All rights reserved.
Figures


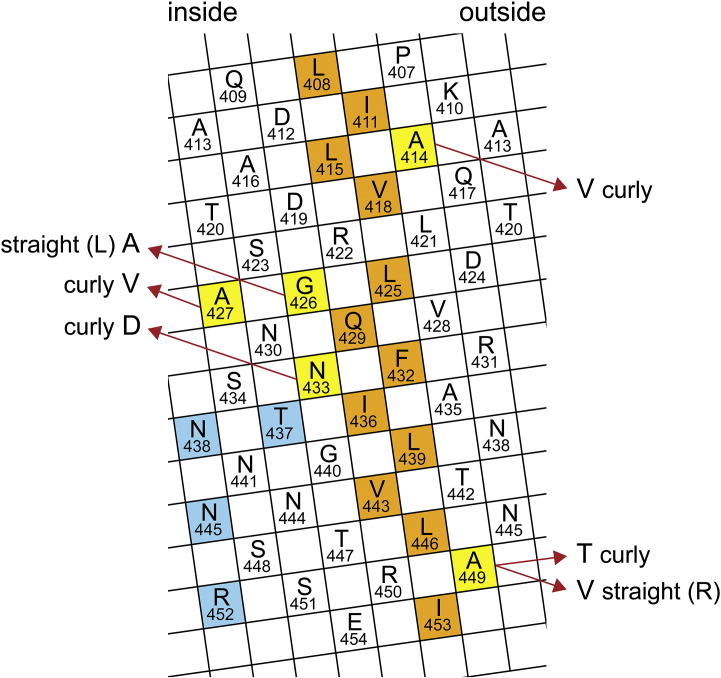
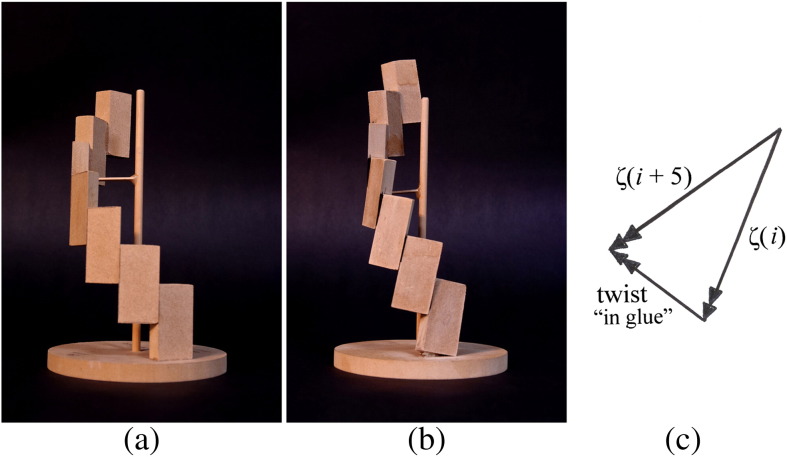
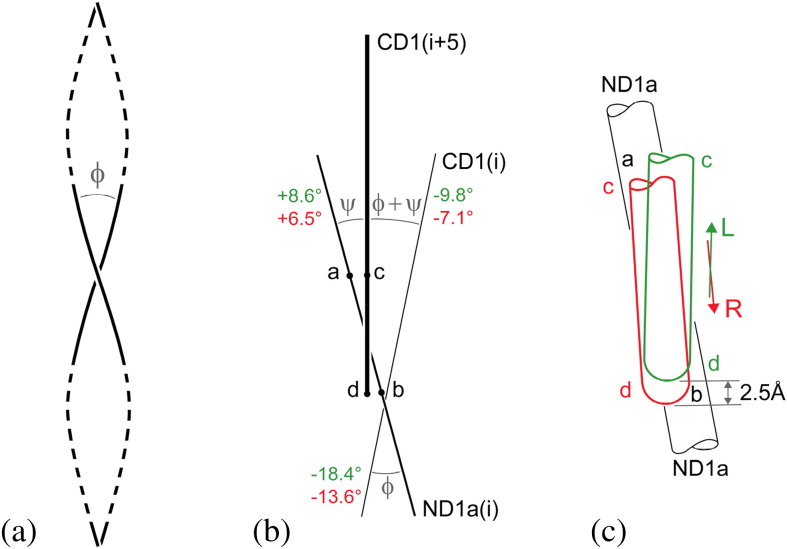
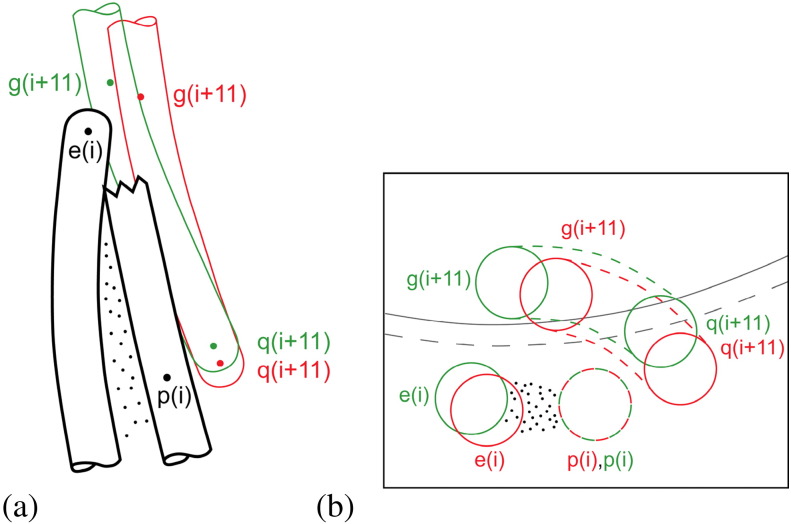
Similar articles
-
Structure and switching of bacterial flagellar filaments studied by X-ray fiber diffraction.Nat Struct Biol. 1998 Feb;5(2):125-32. doi: 10.1038/nsb0298-125. Nat Struct Biol. 1998. PMID: 9461078
-
Crosslinked flagella as a stabilized vaccine adjuvant scaffold.BMC Biotechnol. 2019 Jul 18;19(1):48. doi: 10.1186/s12896-019-0545-3. BMC Biotechnol. 2019. PMID: 31319823 Free PMC article.
-
Structure of the bacterial flagellar protofilament and implications for a switch for supercoiling.Nature. 2001 Mar 15;410(6826):331-7. doi: 10.1038/35066504. Nature. 2001. PMID: 11268201
-
Construction of bacterial flagellar filaments, and aspects of their conversion to different helical forms.Symp Soc Exp Biol. 1982;35:33-51. Symp Soc Exp Biol. 1982. PMID: 6764043 Review.
-
Spinning tails.Curr Opin Struct Biol. 1995 Apr;5(2):187-93. doi: 10.1016/0959-440x(95)80074-3. Curr Opin Struct Biol. 1995. PMID: 7648320 Review.
Cited by
-
The coiled-coil domain of glycosomal membrane-associated Leishmania donovani PEX14: cloning, overexpression, purification and preliminary crystallographic analysis.Acta Crystallogr F Struct Biol Commun. 2020 Oct 1;76(Pt 10):464-468. doi: 10.1107/S2053230X20011127. Epub 2020 Sep 15. Acta Crystallogr F Struct Biol Commun. 2020. PMID: 33006573 Free PMC article.
-
An asymmetric sheath controls flagellar supercoiling and motility in the leptospira spirochete.Elife. 2020 Mar 11;9:e53672. doi: 10.7554/eLife.53672. Elife. 2020. PMID: 32157997 Free PMC article.
-
Flagellin outer domain dimerization modulates motility in pathogenic and soil bacteria from viscous environments.Nat Commun. 2022 Mar 17;13(1):1422. doi: 10.1038/s41467-022-29069-y. Nat Commun. 2022. PMID: 35301306 Free PMC article.
-
The swimming of a deforming helix.Eur Phys J E Soft Matter. 2018 Oct 11;41(10):119. doi: 10.1140/epje/i2018-11728-2. Eur Phys J E Soft Matter. 2018. PMID: 30302671
-
Transcriptional Control of the Lateral-Flagellar Genes of Bradyrhizobium diazoefficiens.J Bacteriol. 2017 Jul 11;199(15):e00253-17. doi: 10.1128/JB.00253-17. Print 2017 Aug 1. J Bacteriol. 2017. PMID: 28533217 Free PMC article.
References
-
- Asakura S. Polymerization of flagellin and polymorphism of flagella. Adv. Biophys. J. 1970;1:99–155. - PubMed
-
- Berg H.C., Anderson R.A. Bacteria swim by rotating their flagellar filaments. Nature. 1973;245:380–382. - PubMed
-
- Larsen S.H., Reader R.W., Kort E.N., Tso W.W., Adler J. Change in direction of flagellar rotation is the basis of the chemotactic response in Escherichia coli. Nature. 1974;249:74–77. - PubMed
-
- Macnab R.M., Ornston M.K. Normal-to-curly flagellar transitions and their role in bacterial tumbling: stabilization of an alternative quaternary structure by mechanical force. J. Mol. Biol. 1977;112:1–30. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
