

ADAR1 promotes malignant progenitor reprogramming in chronic myeloid leukemia
- PMID: 23275297
- PMCID: PMC3549099
- DOI: 10.1073/pnas.1213021110
ADAR1 promotes malignant progenitor reprogramming in chronic myeloid leukemia
Abstract
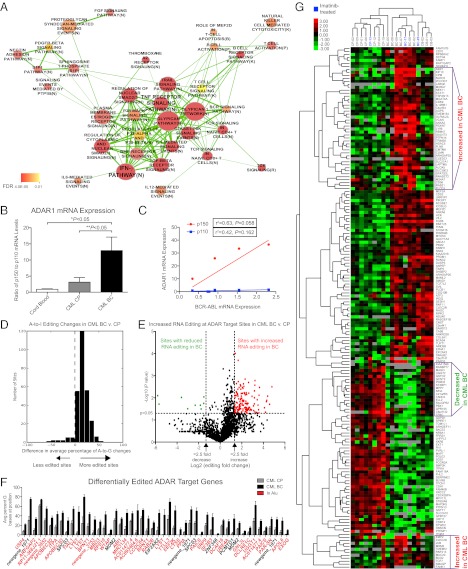
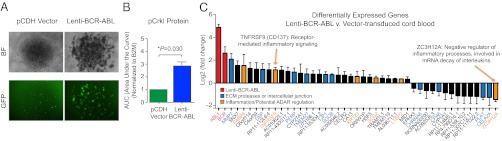
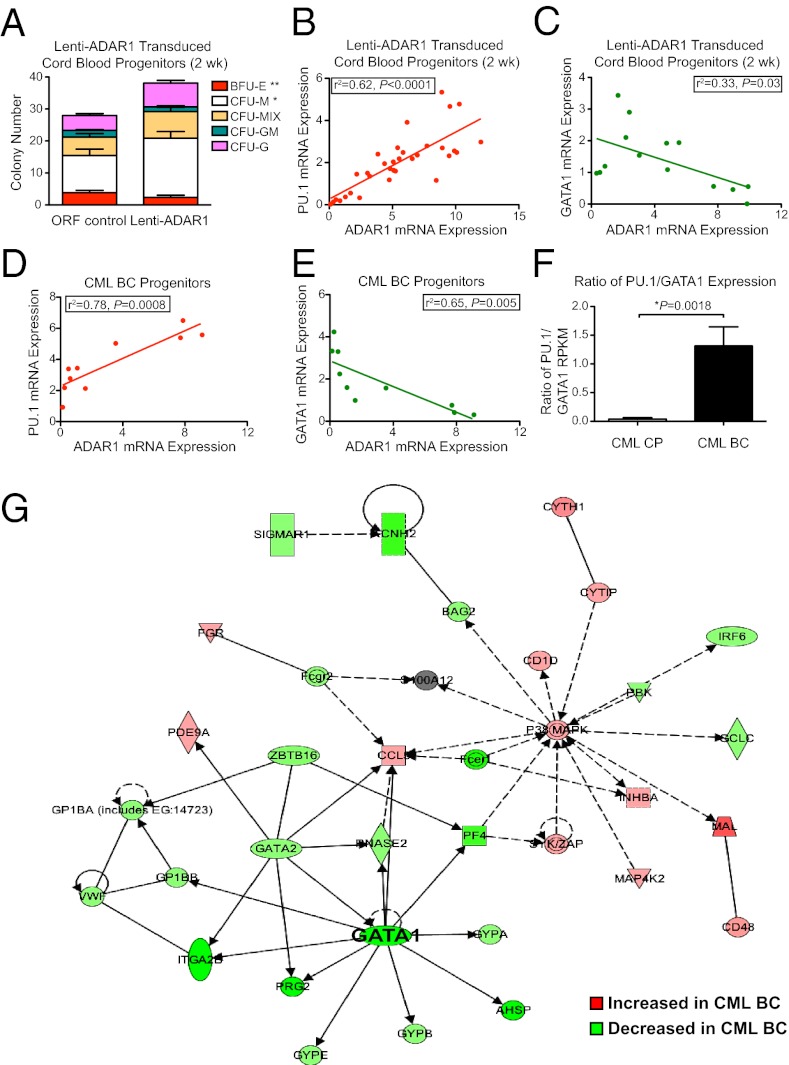
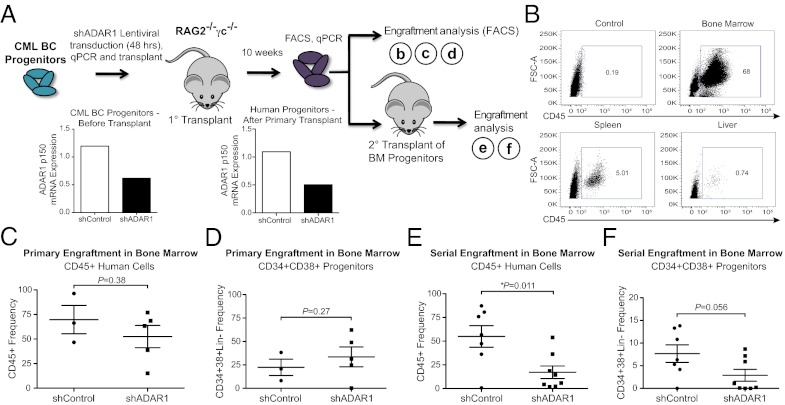
The molecular etiology of human progenitor reprogramming into self-renewing leukemia stem cells (LSC) has remained elusive. Although DNA sequencing has uncovered spliceosome gene mutations that promote alternative splicing and portend leukemic transformation, isoform diversity also may be generated by RNA editing mediated by adenosine deaminase acting on RNA (ADAR) enzymes that regulate stem cell maintenance. In this study, whole-transcriptome sequencing of normal, chronic phase, and serially transplantable blast crisis chronic myeloid leukemia (CML) progenitors revealed increased IFN-γ pathway gene expression in concert with BCR-ABL amplification, enhanced expression of the IFN-responsive ADAR1 p150 isoform, and a propensity for increased adenosine-to-inosine RNA editing during CML progression. Lentiviral overexpression experiments demonstrate that ADAR1 p150 promotes expression of the myeloid transcription factor PU.1 and induces malignant reprogramming of myeloid progenitors. Moreover, enforced ADAR1 p150 expression was associated with production of a misspliced form of GSK3β implicated in LSC self-renewal. Finally, functional serial transplantation and shRNA studies demonstrate that ADAR1 knockdown impaired in vivo self-renewal capacity of blast crisis CML progenitors. Together these data provide a compelling rationale for developing ADAR1-based LSC detection and eradication strategies.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Jamieson C. The MLLgnant consequences of reverting to an embryonic transcriptional program. Cell Stem Cell. 2009;4(2):97–98. - PubMed
-
- Eppert K, et al. Stem cell gene expression programs influence clinical outcome in human leukemia. Nat Med. 2011;17(9):1086–1093. - PubMed
-
- Ben-Neriah Y, Daley GQ, Mes-Masson AM, Witte ON, Baltimore D. The chronic myelogenous leukemia-specific P210 protein is the product of the bcr/abl hybrid gene. Science. 1986;233(4760):212–214. - PubMed
-
- Rowley JD. Letter: A new consistent chromosomal abnormality in chronic myelogenous leukaemia identified by quinacrine fluorescence and Giemsa staining. Nature. 1973;243(5405):290–293. - PubMed
-
- Shtivelman E, Lifshitz B, Gale RP, Canaani E. Fused transcript of abl and bcr genes in chronic myelogenous leukaemia. Nature. 1985;315(6020):550–554. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
