

Lipid droplets and peroxisomes: key players in cellular lipid homeostasis or a matter of fat--store 'em up or burn 'em down
- PMID: 23275493
- PMCID: PMC3527239
- DOI: 10.1534/genetics.112.143362
Lipid droplets and peroxisomes: key players in cellular lipid homeostasis or a matter of fat--store 'em up or burn 'em down
Abstract
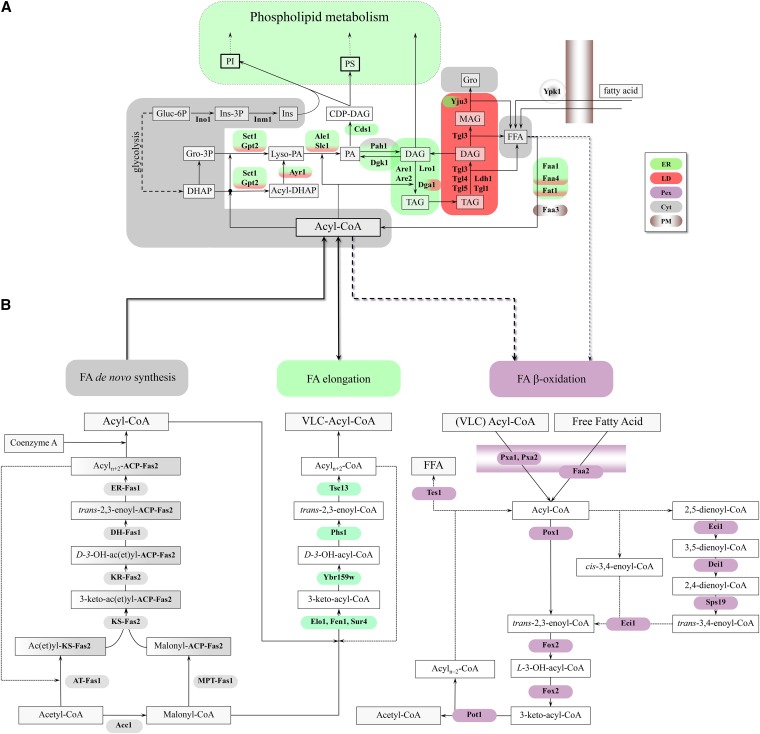
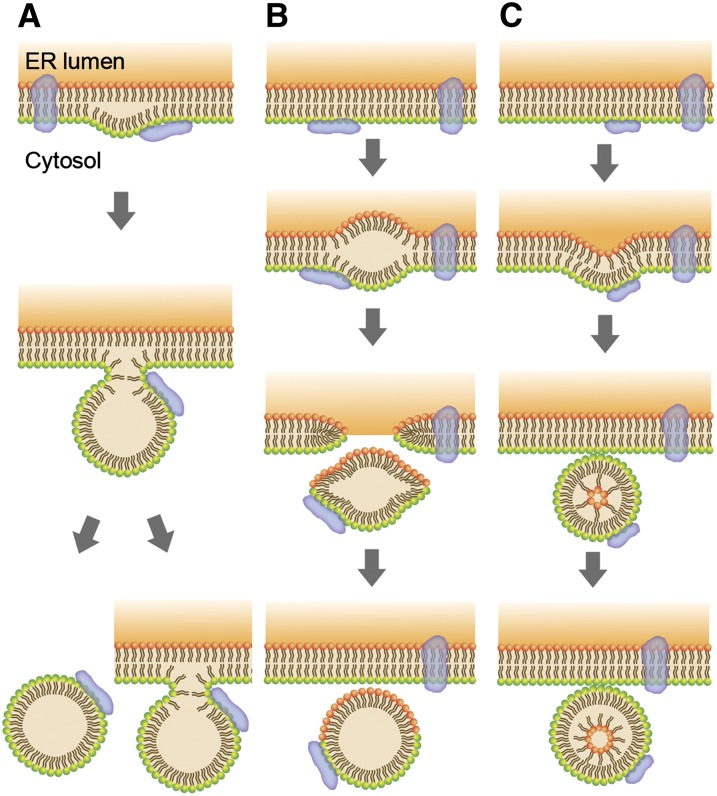
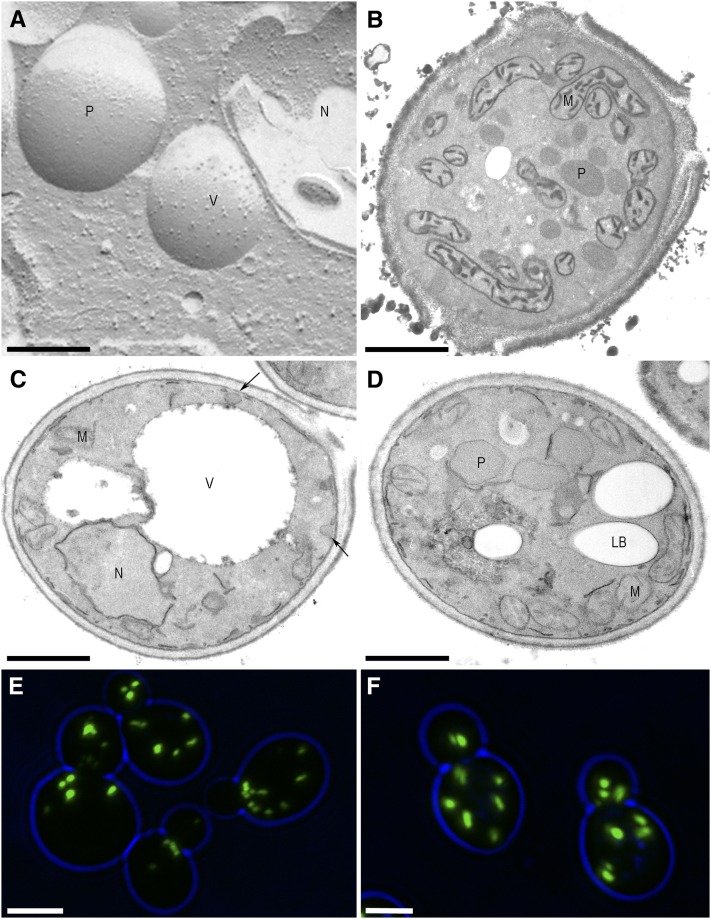
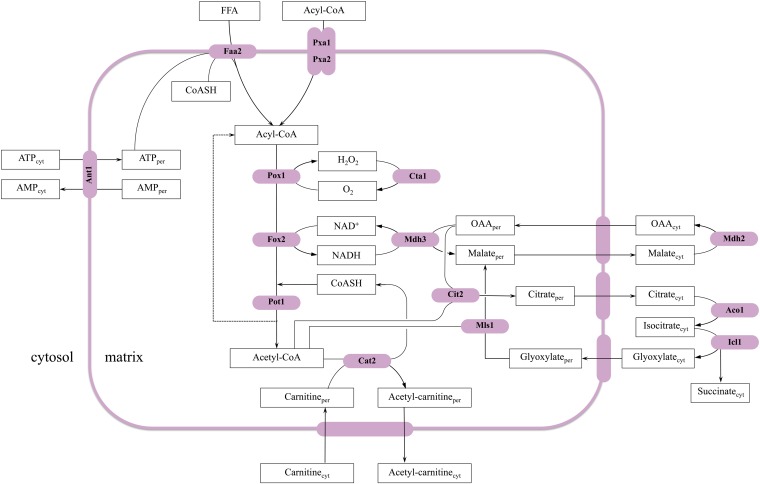
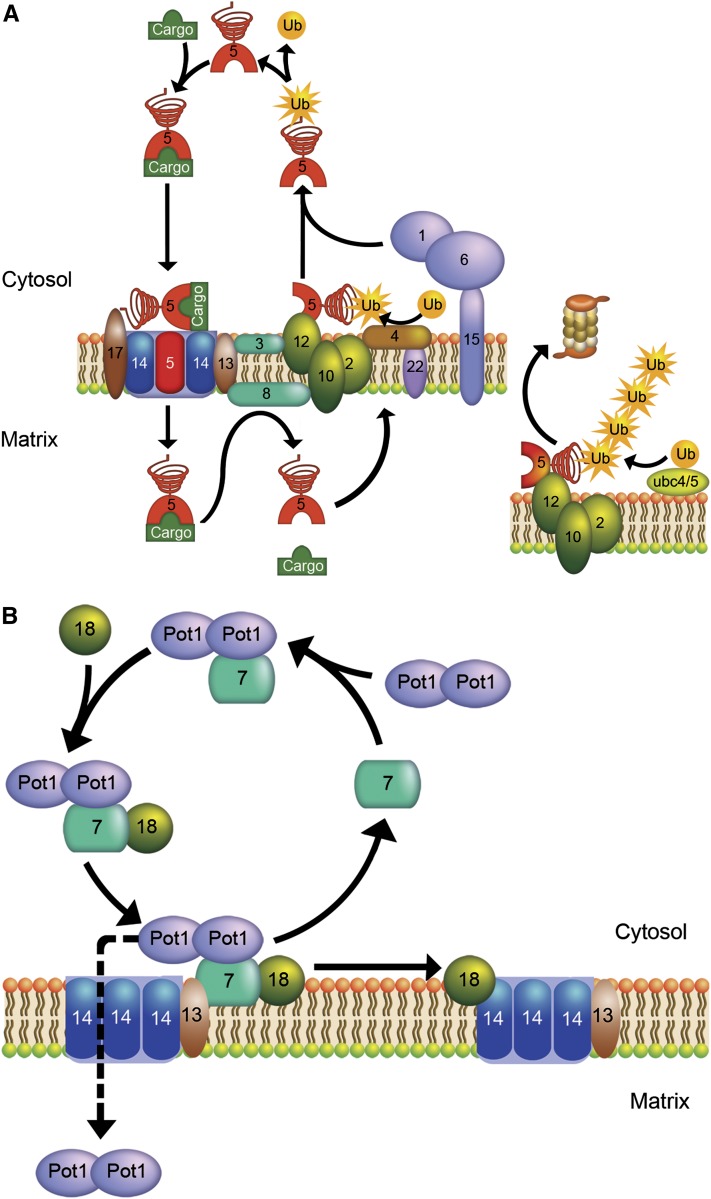
Lipid droplets (LDs) and peroxisomes are central players in cellular lipid homeostasis: some of their main functions are to control the metabolic flux and availability of fatty acids (LDs and peroxisomes) as well as of sterols (LDs). Both fatty acids and sterols serve multiple functions in the cell-as membrane stabilizers affecting membrane fluidity, as crucial structural elements of membrane-forming phospholipids and sphingolipids, as protein modifiers and signaling molecules, and last but not least, as a rich carbon and energy source. In addition, peroxisomes harbor enzymes of the malic acid shunt, which is indispensable to regenerate oxaloacetate for gluconeogenesis, thus allowing yeast cells to generate sugars from fatty acids or nonfermentable carbon sources. Therefore, failure of LD and peroxisome biogenesis and function are likely to lead to deregulated lipid fluxes and disrupted energy homeostasis with detrimental consequences for the cell. These pathological consequences of LD and peroxisome failure have indeed sparked great biomedical interest in understanding the biogenesis of these organelles, their functional roles in lipid homeostasis, interaction with cellular metabolism and other organelles, as well as their regulation, turnover, and inheritance. These questions are particularly burning in view of the pandemic development of lipid-associated disorders worldwide.
Figures

References
-
- AbdelRaheim S. R., Cartwright J. L., Gasmi L., McLennan A. G., 2001. The NADH diphosphatase encoded by the Saccharomyces cerevisiae NPY1 nudix hydrolase gene is located in peroxisomes. Arch. Biochem. Biophys. 388: 18–24 - PubMed
-
- Agarwal A. K., Garg A., 2003. Congenital generalized lipodystrophy: significance of triglyceride biosynthetic pathways. Trends Endocrinol. Metab. 14: 214–221 - PubMed
-
- Agarwal A. K., Barnes R. I., Garg A., 2004. Genetic basis of congenital generalized lipodystrophy. Int. J. Obes. Relat. Metab. Disord. 28: 336–339 - PubMed
-
- Agne B., Meindl N. M., Niederhoff K., Einwachter H., Rehling P., et al. , 2003. Pex8p: an intraperoxisomal organizer of the peroxisomal import machinery. Mol. Cell 11: 635–646 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
