

De novo design of a synthetic riboswitch that regulates transcription termination
- PMID: 23275562
- PMCID: PMC3575828
- DOI: 10.1093/nar/gks1330
De novo design of a synthetic riboswitch that regulates transcription termination
Abstract
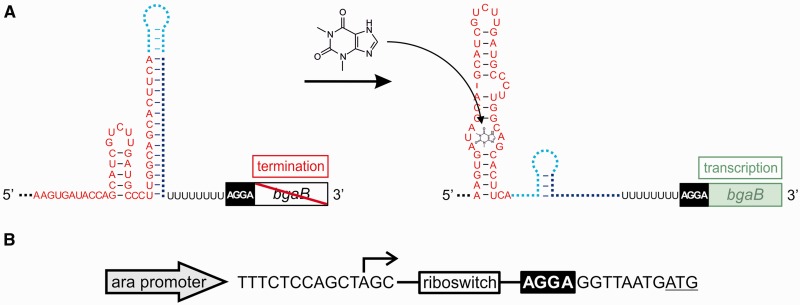
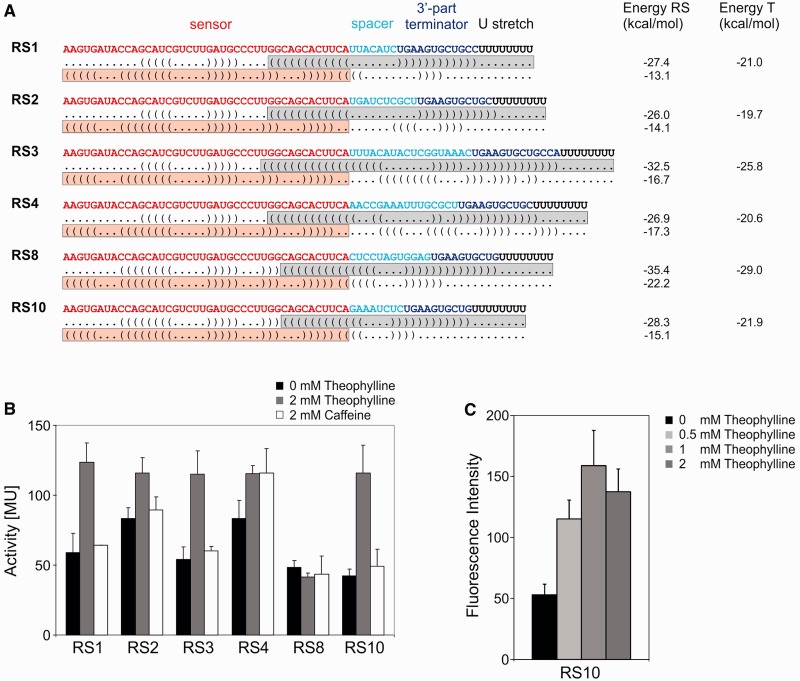
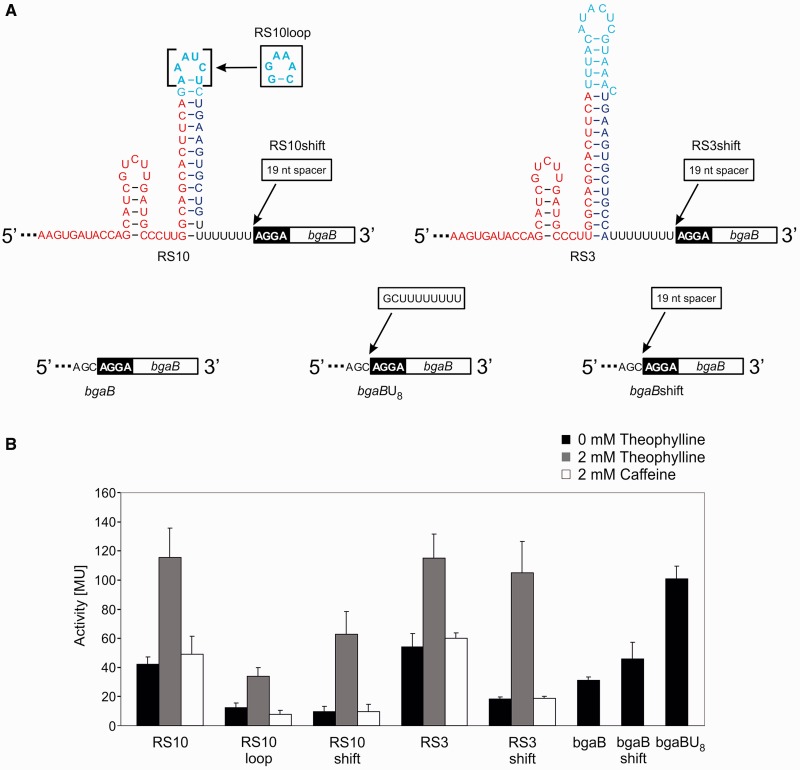
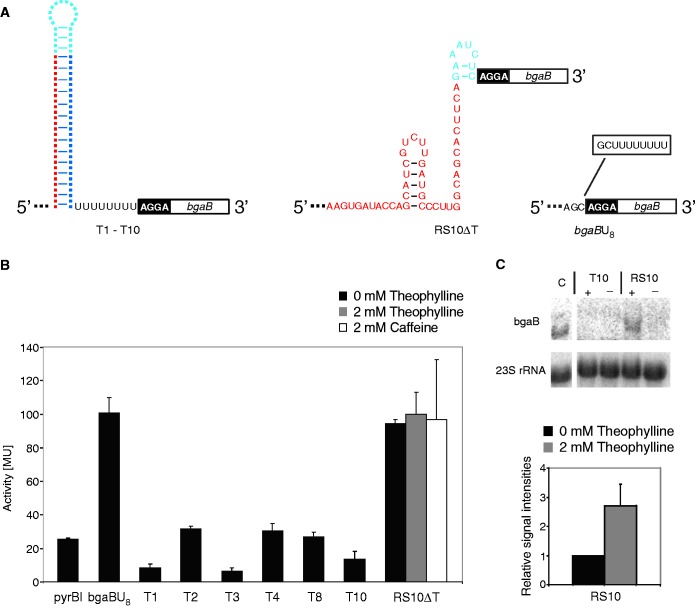
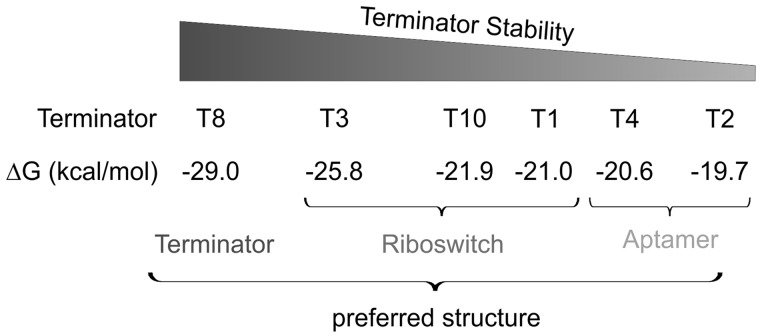
Riboswitches are regulatory RNA elements typically located in the 5'-untranslated region of certain mRNAs and control gene expression at the level of transcription or translation. These elements consist of a sensor and an adjacent actuator domain. The sensor usually is an aptamer that specifically interacts with a ligand. The actuator contains an intrinsic terminator or a ribosomal binding site for transcriptional or translational regulation, respectively. Ligand binding leads to structural rearrangements of the riboswitch and to presentation or masking of these regulatory elements. Based on this modular organization, riboswitches are an ideal target for constructing synthetic regulatory systems for gene expression. Although riboswitches for translational control have been designed successfully, attempts to construct synthetic elements regulating transcription have failed so far. Here, we present an in silico pipeline for the rational design of synthetic riboswitches that regulate gene expression at the transcriptional level. Using the well-characterized theophylline aptamer as sensor, we designed the actuator part as RNA sequences that can fold into functional intrinsic terminator structures. In the biochemical characterization, several of the designed constructs show ligand-dependent control of gene expression in Escherichia coli, demonstrating that it is possible to engineer riboswitches not only for translational but also for transcriptional regulation.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
