

Developmental changes in the cochlear hair cell mechanotransducer channel and their regulation by transmembrane channel-like proteins
- PMID: 23277480
- PMCID: PMC3536526
- DOI: 10.1085/jgp.201210913
Developmental changes in the cochlear hair cell mechanotransducer channel and their regulation by transmembrane channel-like proteins
Abstract
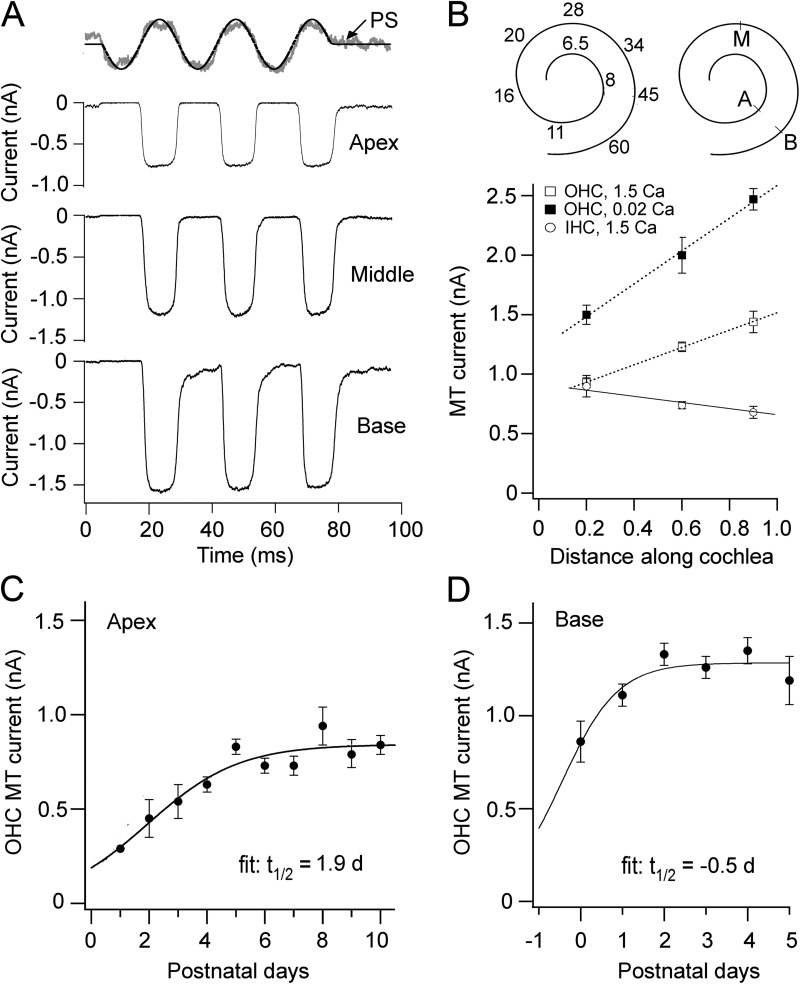
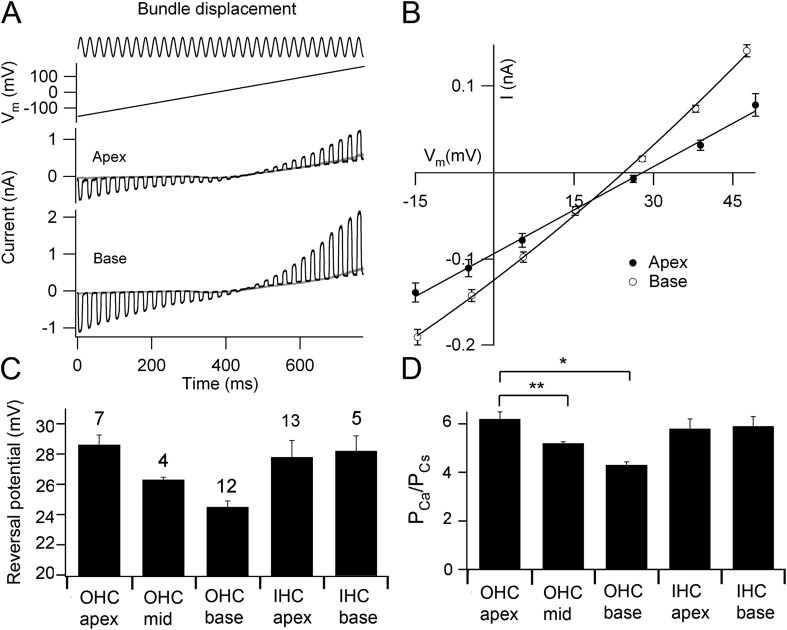
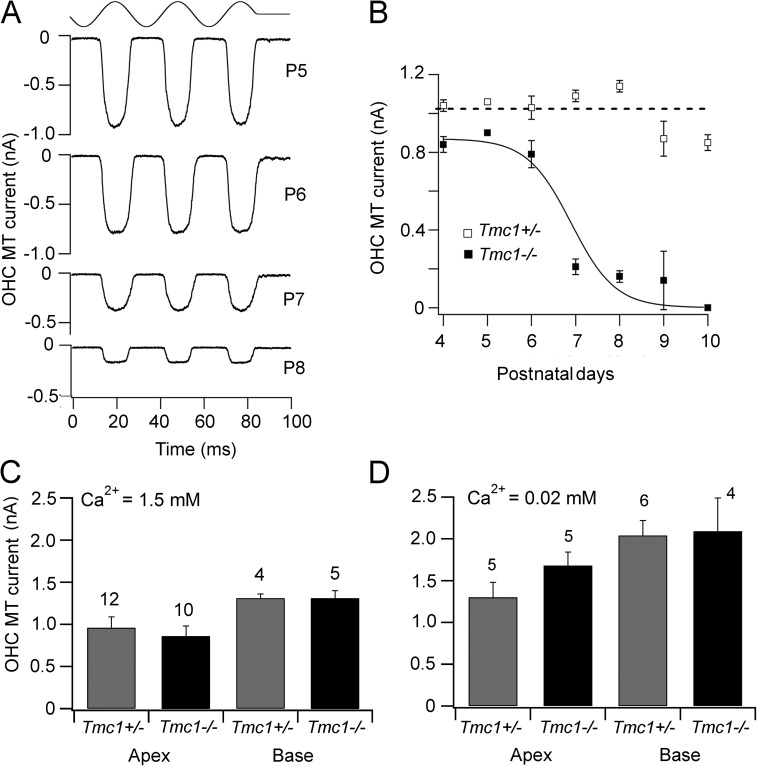
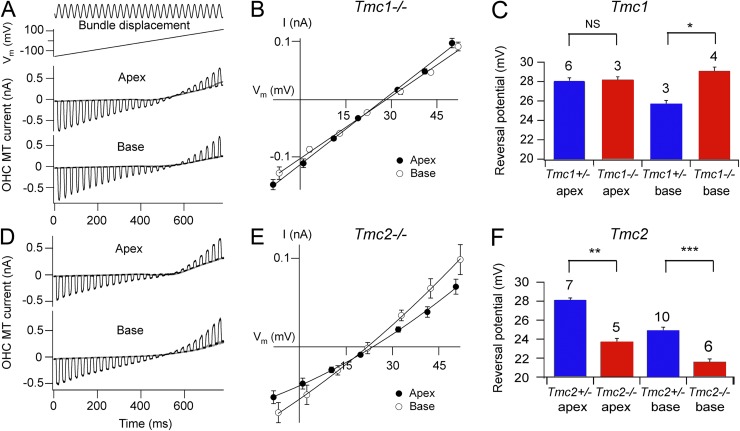
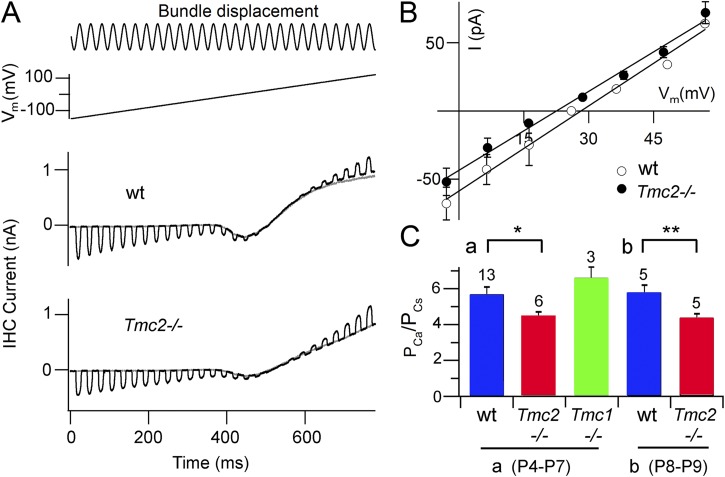
Vibration of the stereociliary bundles activates calcium-permeable mechanotransducer (MT) channels to initiate sound detection in cochlear hair cells. Different regions of the cochlea respond preferentially to different acoustic frequencies, with variation in the unitary conductance of the MT channels contributing to this tonotopic organization. Although the molecular identity of the MT channel remains uncertain, two members of the transmembrane channel-like family, Tmc1 and Tmc2, are crucial to hair cell mechanotransduction. We measured MT channel current amplitude and Ca(2+) permeability along the cochlea's longitudinal (tonotopic) axis during postnatal development of wild-type mice and mice lacking Tmc1 (Tmc1-/-) or Tmc2 (Tmc2-/-). In wild-type mice older than postnatal day (P) 4, MT current amplitude increased ~1.5-fold from cochlear apex to base in outer hair cells (OHCs) but showed little change in inner hair cells (IHCs), a pattern apparent in mutant mice during the first postnatal week. After P7, the OHC MT current in Tmc1-/- (dn) mice declined to zero, consistent with their deafness phenotype. In wild-type mice before P6, the relative Ca(2+) permeability, P(Ca), of the OHC MT channel decreased from cochlear apex to base. This gradient in P(Ca) was not apparent in IHCs and disappeared after P7 in OHCs. In Tmc1-/- mice, P(Ca) in basal OHCs was larger than that in wild-type mice (to equal that of apical OHCs), whereas in Tmc2-/-, P(Ca) in apical and basal OHCs and IHCs was decreased compared with that in wild-type mice. We postulate that differences in Ca(2+) permeability reflect different subunit compositions of the MT channel determined by expression of Tmc1 and Tmc2, with the latter conferring higher P(Ca) in IHCs and immature apical OHCs. Changes in P(Ca) with maturation are consistent with a developmental decrease in abundance of Tmc2 in OHCs but not in IHCs.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
