

Essential role of postsynaptic NMDA receptors in developmental refinement of excitatory synapses
- PMID: 23277569
- PMCID: PMC3549111
- DOI: 10.1073/pnas.1212971110
Essential role of postsynaptic NMDA receptors in developmental refinement of excitatory synapses
Abstract
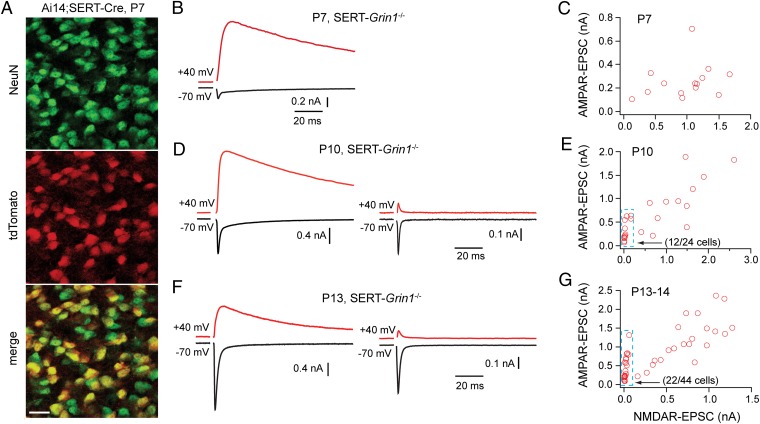
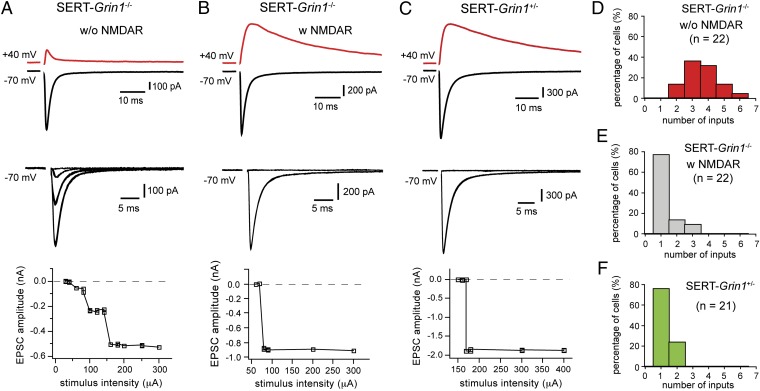
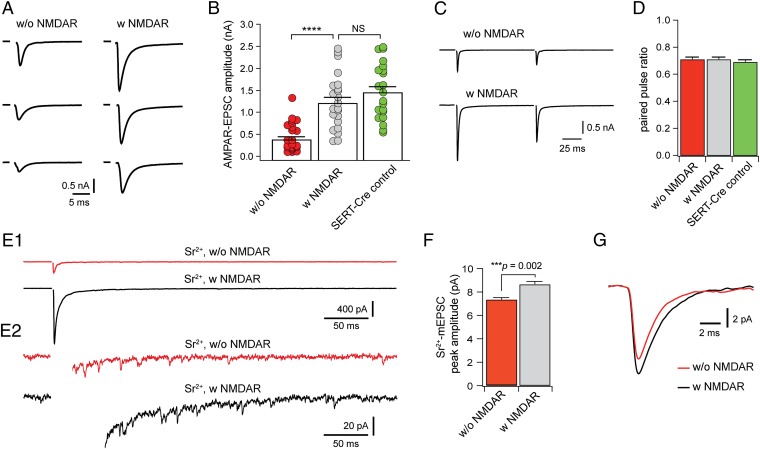
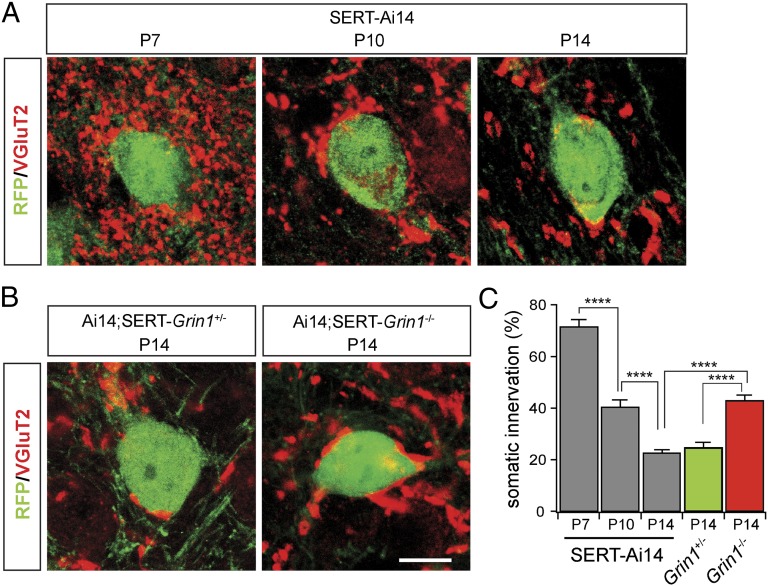
Neurons in the brains of newborns are usually connected with many other neurons through weak synapses. This early pattern of connectivity is refined through pruning of many immature connections and strengthening of the remaining ones. NMDA receptors (NMDARs) are essential for the development of excitatory synapses, but their role in synaptic refinement is controversial. Although chronic application of blockers or global knockdown of NMDARs disrupts developmental refinement in many parts of the brain, the ubiquitous presence of NMDARs makes it difficult to dissociate direct effects from indirect ones. We addressed this question in the thalamus by using genetic mosaic deletion of NMDARs. We demonstrate that pruning and strengthening of immature synapses are blocked in neurons without NMDARs, but occur normally in neighboring neurons with NMDARs. Our data support a model in which activation of NMDARs in postsynaptic neurons initiates synaptic refinement.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
High Salt Intake Recruits Tonic Activation of NR2D Subunit-Containing Extrasynaptic NMDARs in Vasopressin Neurons.J Neurosci. 2021 Feb 10;41(6):1145-1156. doi: 10.1523/JNEUROSCI.1742-20.2020. Epub 2020 Dec 10. J Neurosci. 2021. PMID: 33303677 Free PMC article.
-
NMDA receptor-dependent pattern transfer from afferents to postsynaptic cells and dendritic differentiation in the barrel cortex.Mol Cell Neurosci. 2002 Nov;21(3):477-92. doi: 10.1006/mcne.2002.1195. Mol Cell Neurosci. 2002. PMID: 12498788 Free PMC article.
-
Developmental decrease in NMDA receptor desensitization associated with shift to synapse and interaction with postsynaptic density-95.J Neurosci. 2003 Dec 3;23(35):11244-54. doi: 10.1523/JNEUROSCI.23-35-11244.2003. J Neurosci. 2003. PMID: 14657184 Free PMC article.
-
Making of a Synapse: Recurrent Roles of Drebrin A at Excitatory Synapses Throughout Life.Adv Exp Med Biol. 2017;1006:119-139. doi: 10.1007/978-4-431-56550-5_8. Adv Exp Med Biol. 2017. PMID: 28865018 Review.
-
Hippocampal long-term synaptic plasticity and signal amplification of NMDA receptors.Crit Rev Neurobiol. 2006;18(1-2):71-84. doi: 10.1615/critrevneurobiol.v18.i1-2.80. Crit Rev Neurobiol. 2006. PMID: 17725510 Review.
Cited by
-
Developmental Impairments of Synaptic Refinement in the Thalamus of a Mouse Model of Fragile X Syndrome.Neurosci Bull. 2024 Apr;40(4):439-450. doi: 10.1007/s12264-023-01142-6. Epub 2023 Nov 28. Neurosci Bull. 2024. PMID: 38015349 Free PMC article.
-
GABA-A receptor-dependent mechanisms prevent excessive spine elimination during postnatal maturation of the mouse cortex in vivo.FEBS Lett. 2014 Dec 20;588(24):4551-60. doi: 10.1016/j.febslet.2014.10.023. Epub 2014 Nov 4. FEBS Lett. 2014. PMID: 25447527 Free PMC article.
-
Brain injury in premature neonates: A primary cerebral dysmaturation disorder?Ann Neurol. 2014 Apr;75(4):469-86. doi: 10.1002/ana.24132. Epub 2014 Apr 14. Ann Neurol. 2014. PMID: 24615937 Free PMC article.
-
The Implication of Glial Metabotropic Glutamate Receptors in Alzheimer's Disease.Curr Neuropharmacol. 2023;21(2):164-182. doi: 10.2174/1570159X20666211223140303. Curr Neuropharmacol. 2023. PMID: 34951388 Free PMC article. Review.
-
Prenatal cerebral ischemia triggers dysmaturation of caudate projection neurons.Ann Neurol. 2014 Apr;75(4):508-24. doi: 10.1002/ana.24100. Epub 2014 Mar 13. Ann Neurol. 2014. PMID: 24395459 Free PMC article.
References
-
- Balice-Gordon RJ, Chua CK, Nelson CC, Lichtman JW. Gradual loss of synaptic cartels precedes axon withdrawal at developing neuromuscular junctions. Neuron. 1993;11(5):801–815. - PubMed
-
- Chen C, Regehr WG. Developmental remodeling of the retinogeniculate synapse. Neuron. 2000;28(3):955–966. - PubMed
-
- Hashimoto K, Kano M. Functional differentiation of multiple climbing fiber inputs during synapse elimination in the developing cerebellum. Neuron. 2003;38(5):785–796. - PubMed
-
- Kim G, Kandler K. Elimination and strengthening of glycinergic/GABAergic connections during tonotopic map formation. Nat Neurosci. 2003;6(3):282–290. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
