

Functioning of a metabolic flux sensor in Escherichia coli
- PMID: 23277571
- PMCID: PMC3549114
- DOI: 10.1073/pnas.1202582110
Functioning of a metabolic flux sensor in Escherichia coli
Abstract
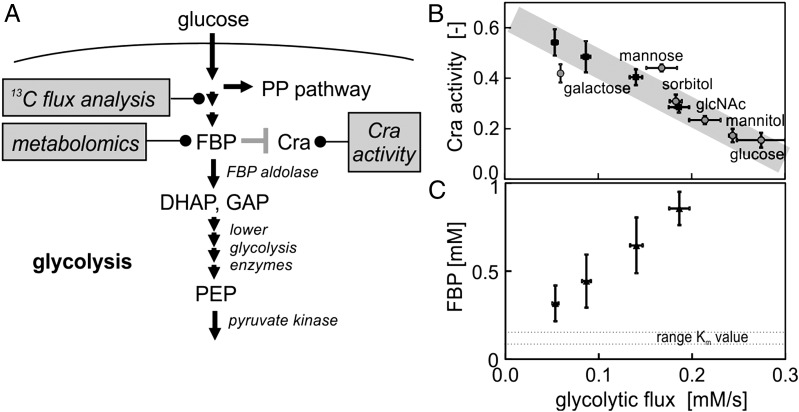
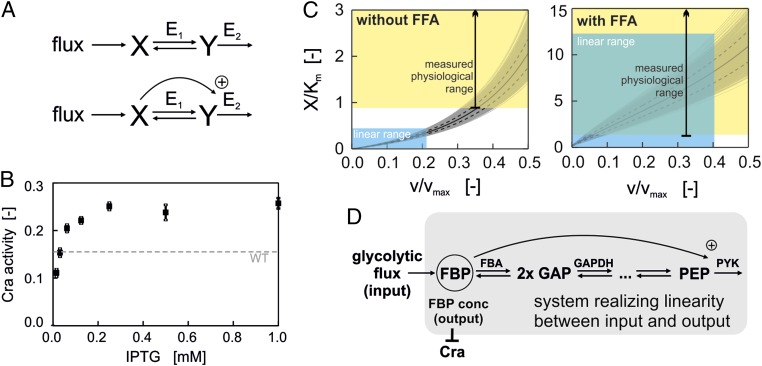
Regulation of metabolic operation in response to extracellular cues is crucial for cells' survival. Next to the canonical nutrient sensors, which measure the concentration of nutrients, recently intracellular "metabolic flux" was proposed as a novel impetus for metabolic regulation. According to this concept, cells would have molecular systems ("flux sensors") in place that regulate metabolism as a function of the actually occurring metabolic fluxes. Although this resembles an appealing concept, we have not had any experimental evidence for the existence of flux sensors and also we have not known how these flux sensors would work in detail. Here, we show experimental evidence that supports the hypothesis that Escherichia coli is indeed able to measure its glycolytic flux and uses this signal for metabolic regulation. Combining experiment and theory, we show how this flux-sensing function could emerge from an aggregate of several molecular mechanisms: First, the system of reactions of lower glycolysis and the feedforward activation of fructose-1,6-bisphosphate on pyruvate kinase translate flux information into the concentration of the metabolite fructose-1,6-bisphosphate. The interaction of this "flux-signaling metabolite" with the transcription factor Cra then leads to flux-dependent regulation. By responding to glycolytic flux, rather than to the concentration of individual carbon sources, the cell may minimize sensing and regulatory expenses.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Martínez-Antonio A, Janga SC, Salgado H, Collado-Vides J. Internal-sensing machinery directs the activity of the regulatory network in Escherichia coli. Trends Microbiol. 2006;14(1):22–27. - PubMed
-
- Sellick CA, Reece RJ. Eukaryotic transcription factors as direct nutrient sensors. Trends Biochem Sci. 2005;30(7):405–412. - PubMed
-
- Holsbeeks I, Lagatie O, Van Nuland A, Van de Velde S, Thevelein JM. The eukaryotic plasma membrane as a nutrient-sensing device. Trends Biochem Sci. 2004;29(10):556–564. - PubMed
-
- Gerosa L, Sauer U. Regulation and control of metabolic fluxes in microbes. Curr Opin Biotechnol. 2011;22(4):566–575. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
