

Energetic cost of protein import across the envelope membranes of chloroplasts
- PMID: 23277572
- PMCID: PMC3549074
- DOI: 10.1073/pnas.1115886110
Energetic cost of protein import across the envelope membranes of chloroplasts
Abstract
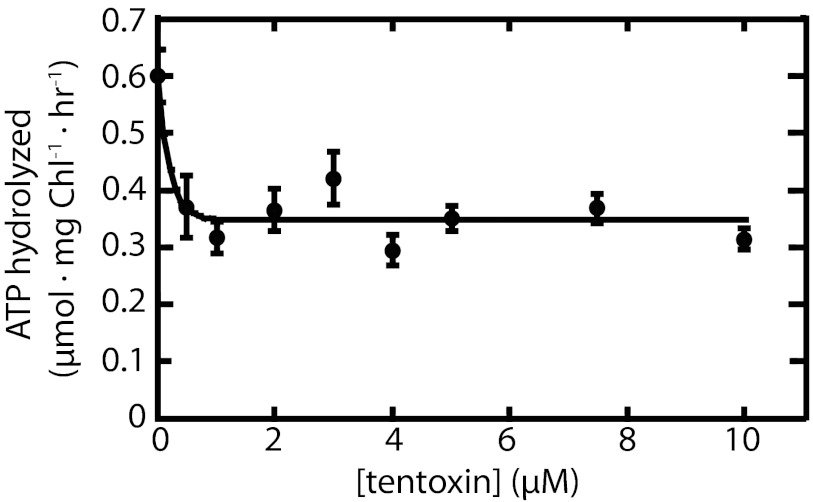
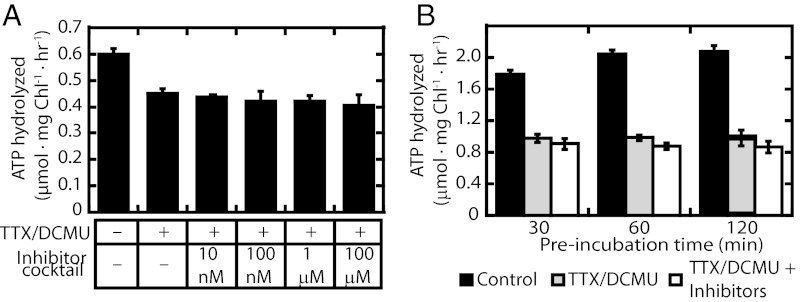
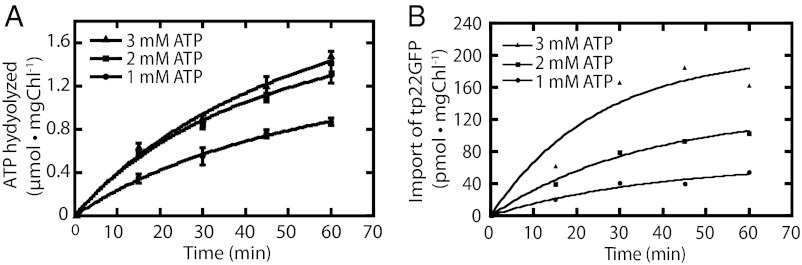
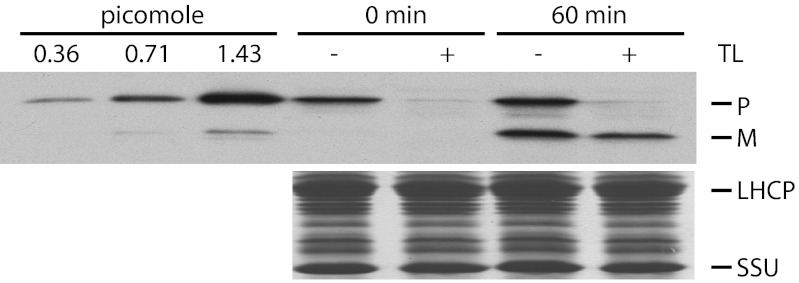
Chloroplasts are the organelles of green plants in which light energy is transduced into chemical energy, forming ATP and reduced carbon compounds upon which all life depends. The expenditure of this energy is one of the central issues of cellular metabolism. Chloroplasts contain ~3,000 proteins, among which less than 100 are typically encoded in the plastid genome. The rest are encoded in the nuclear genome, synthesized in the cytosol, and posttranslationally imported into the organelle in an energy-dependent process. We report here a measurement of the amount of ATP hydrolyzed to import a protein across the chloroplast envelope membranes--only the second complete accounting of the cost in Gibbs free energy of protein transport to be undertaken. Using two different precursors prepared by three distinct techniques, we show that the import of a precursor protein into chloroplasts is accompanied by the hydrolysis of ~650 ATP molecules. This translates to a ΔG(protein) (transport) of some 27,300 kJ/mol protein imported. We estimate that protein import across the plastid envelope membranes consumes ~0.6% of the total light-saturated energy output of the organelle.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Schatz G, Dobberstein B. Common principles of protein translocation across membranes. Science. 1996;271(5255):1519–1526. - PubMed
-
- Mori T, Ishitani R, Tsukazaki T, Nureki O, Sugita Y. Molecular mechanisms underlying the early stage of protein translocation through the Sec translocon. Biochemistry. 2010;49(5):945–950. - PubMed
-
- Driessen AJ, Nouwen N. Protein translocation across the bacterial cytoplasmic membrane. Annu Rev Biochem. 2008;77:643–667. - PubMed
-
- Alder NN, Theg SM. Energy use by biological protein transport pathways. Trends Biochem Sci. 2003;28(8):442–451. - PubMed
-
- Johnson AE, Haigh NG. The ER translocon and retrotranslocation: Is the shift into reverse manual or automatic? Cell. 2000;102(6):709–712. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
