

C/EBP homologous protein (CHOP) contributes to suppression of metabolic genes during endoplasmic reticulum stress in the liver
- PMID: 23281479
- PMCID: PMC3567690
- DOI: 10.1074/jbc.M112.432344
C/EBP homologous protein (CHOP) contributes to suppression of metabolic genes during endoplasmic reticulum stress in the liver
Abstract
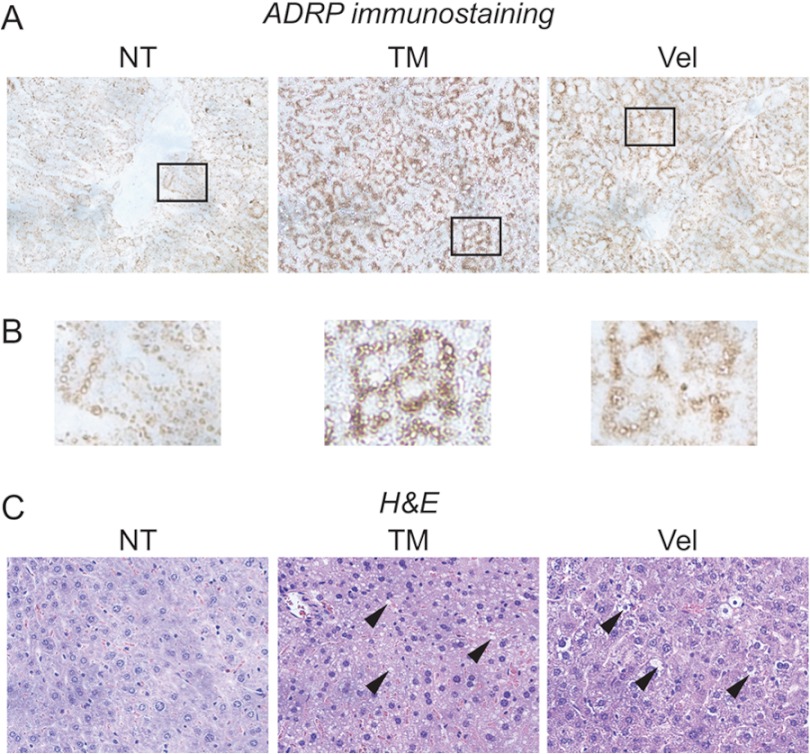
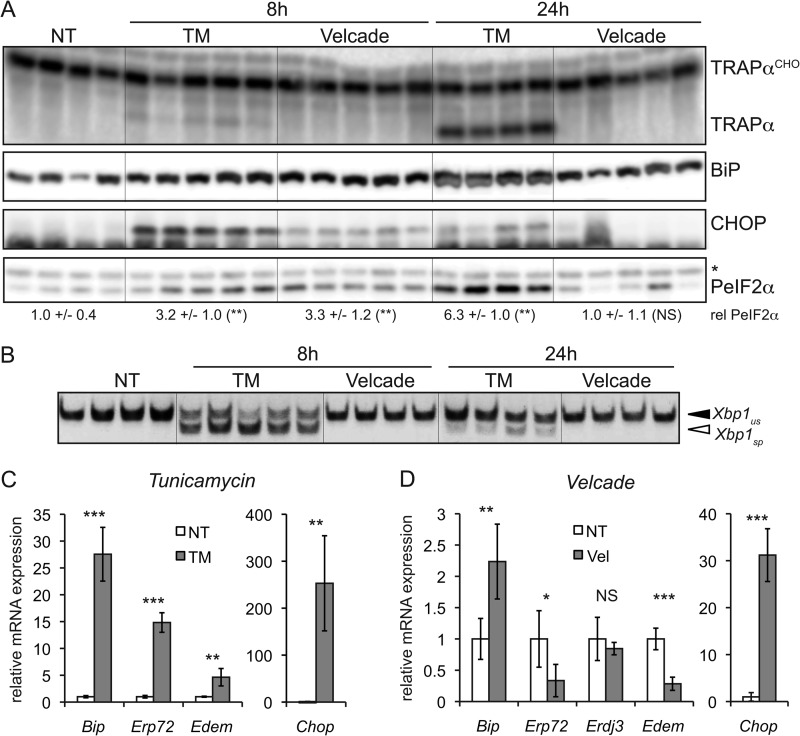
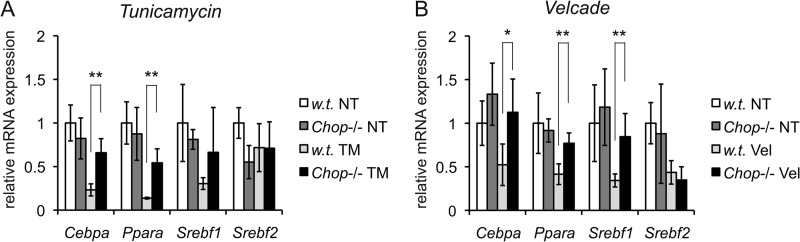
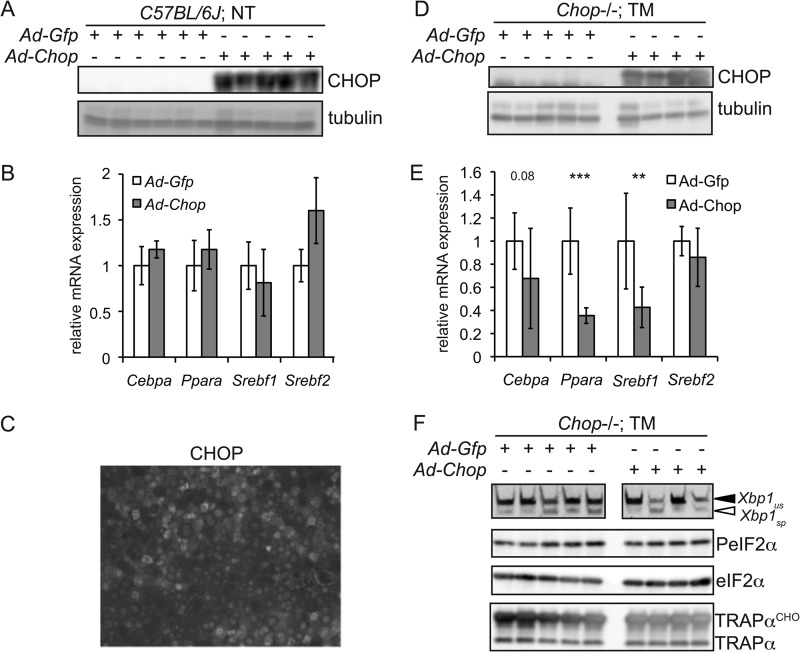
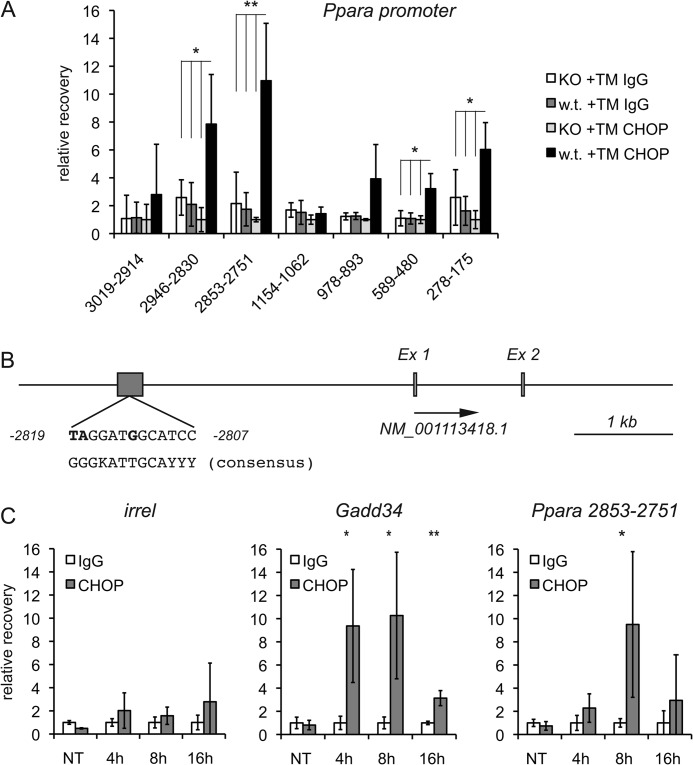
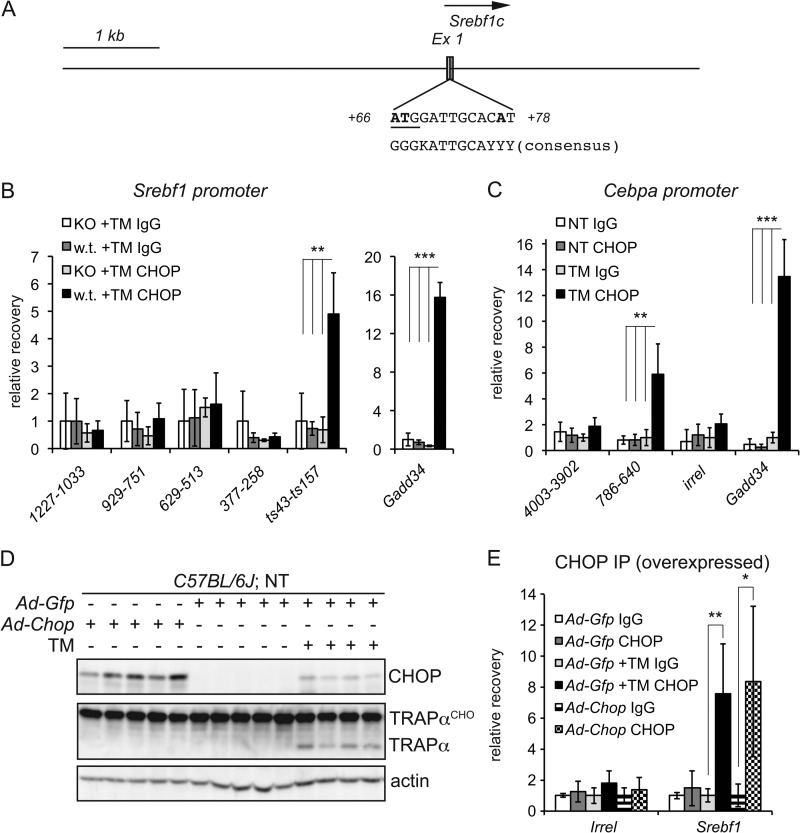
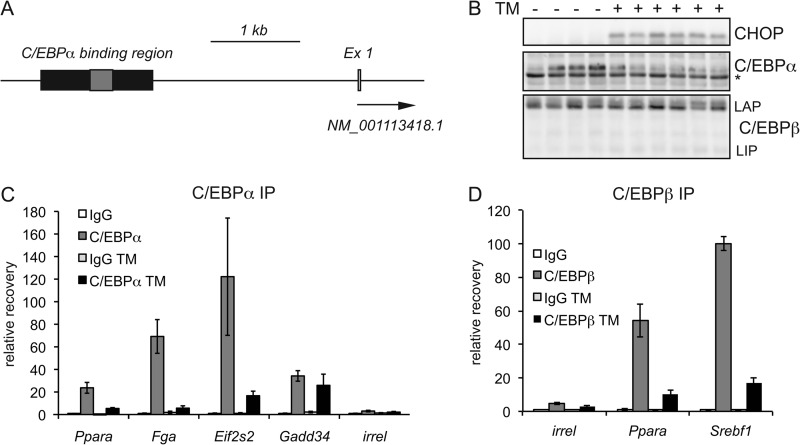
The unfolded protein response (UPR) senses stress in the endoplasmic reticulum (ER) and initiates signal transduction cascades that culminate in changes to gene regulation. Long recognized as a means for improving ER protein folding through up-regulation of ER chaperones, the UPR is increasingly recognized to play a role in the regulation of metabolic pathways. ER stress is clearly connected to altered metabolism in tissues such as the liver, but the mechanisms underlying this connection are only beginning to be elucidated. Here, working exclusively in vivo, we tested the hypothesis that the UPR-regulated CCAAT/enhancer-binding protein (C/EBP) homologous protein (CHOP) participates in the transcriptional regulation of metabolism during hepatic ER stress. We found that metabolic dysregulation was associated with induction of eIF2α signaling and CHOP up-regulation during challenge with tunicamycin or Velcade. CHOP was necessary for suppression of genes encoding the transcriptional master regulators of lipid metabolism: Cebpa, Ppara, and Srebf1. This action of CHOP required a contemporaneous CHOP-independent stress signal. CHOP bound directly to C/EBP-binding regions in the promoters of target genes, whereas binding of C/EBPα and C/EBPβ to the same regions was diminished during ER stress. Our results thus highlight a role for CHOP in the transcriptional regulation of metabolism.
Figures
References
-
- Walter P., Ron D. (2011) The unfolded protein response: from stress pathway to homeostatic regulation. Science 334, 1081–1086 - PubMed
-
- Harding H. P., Novoa I., Zhang Y., Zeng H., Wek R., Schapira M., Ron D. (2000) Regulated translation initiation controls stress-induced gene expression in mammalian cells. Mol. Cell 6, 1099–1108 - PubMed
-
- Harding H. P., Zhang Y., Bertolotti A., Zeng H., Ron D. (2000) Perk is essential for translational regulation and cell survival during the unfolded protein response. Mol. Cell 5, 897–904 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
