

Proteoglycans and their roles in brain cancer
- PMID: 23281850
- PMCID: PMC3644380
- DOI: 10.1111/febs.12109
Proteoglycans and their roles in brain cancer
Abstract
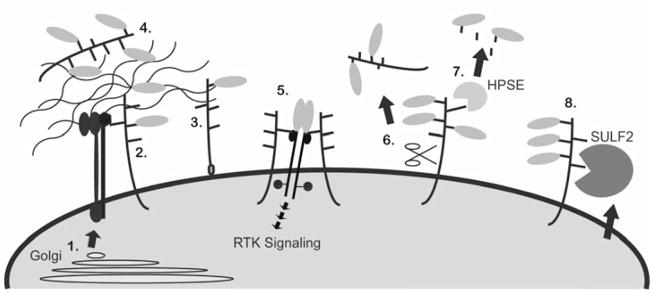
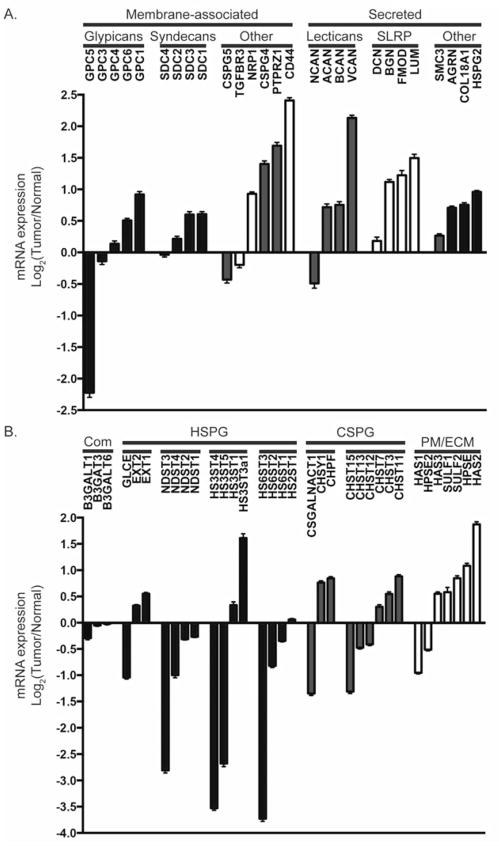
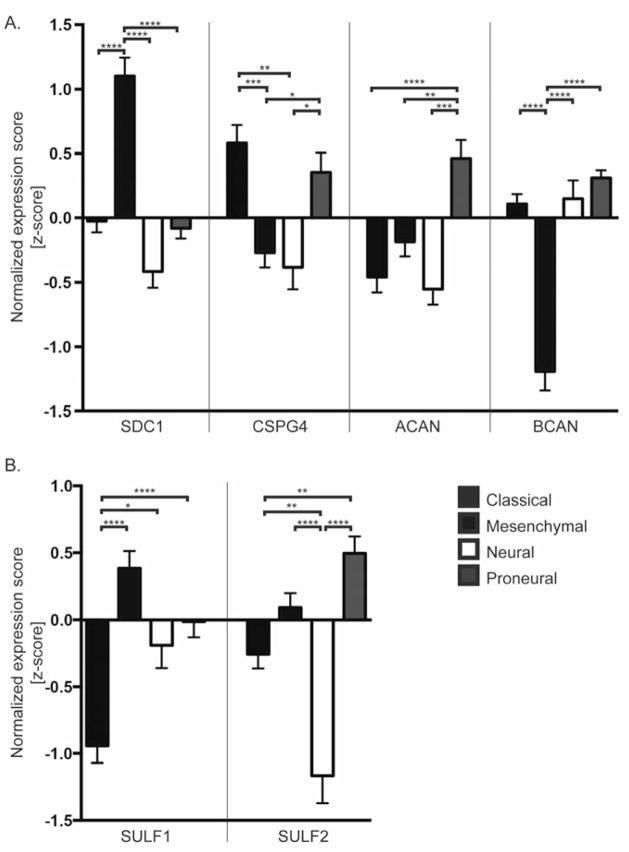
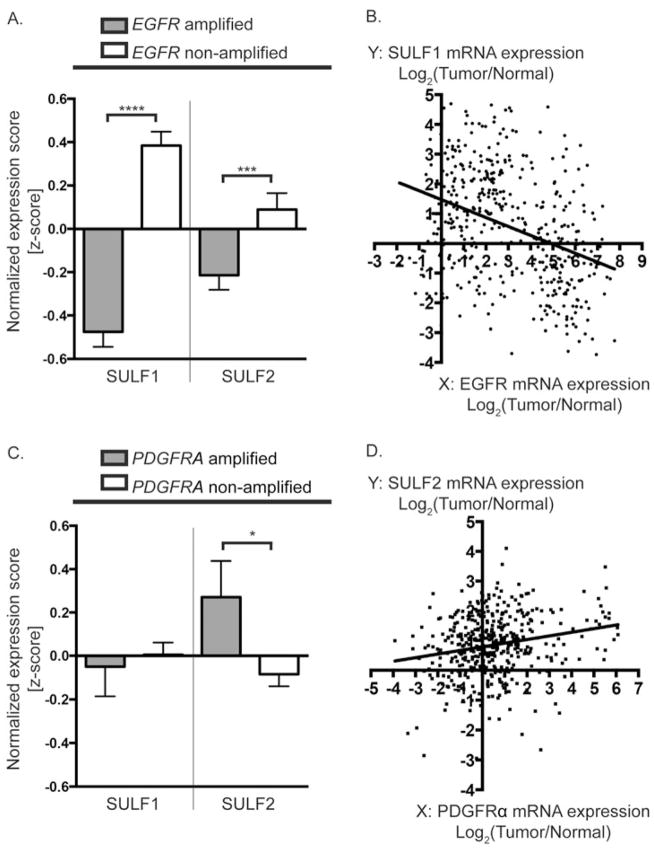
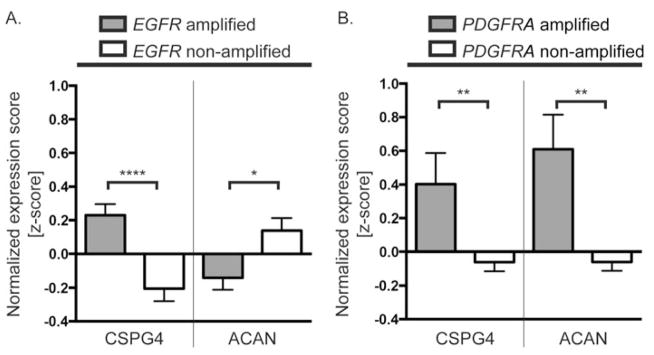
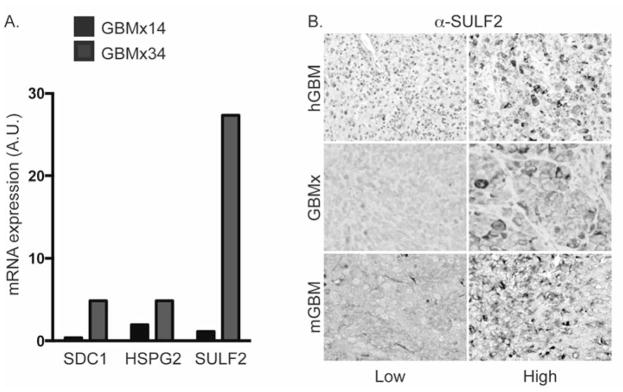
Glioblastoma, a malignant brain cancer, is characterized by abnormal activation of receptor tyrosine kinase signalling pathways and a poor prognosis. Extracellular proteoglycans, including heparan sulfate and chondroitin sulfate, play critical roles in the regulation of cell signalling and migration via interactions with extracellular ligands, growth factor receptors and extracellular matrix components, as well as intracellular enzymes and structural proteins. In cancer, proteoglycans help drive multiple oncogenic pathways in tumour cells and promote critical tumour-microenvironment interactions. In the present review, we summarize the evidence for proteoglycan function in gliomagenesis and examine the expression of proteoglycans and their modifying enzymes in human glioblastoma using data obtained from The Cancer Genome Atlas (http://cancergenome.nih.gov/). Furthermore, we demonstrate an association between specific proteoglycan alterations and changes in receptor tyrosine kinases. Based on these data, we propose a model in which proteoglycans and their modifying enzymes promote receptor tyrosine kinase signalling and progression in glioblastoma, and we suggest that cancer-associated proteoglycans are promising biomarkers for disease and therapeutic targets.
© 2013 The Authors Journal compilation © 2013 FEBS.
Conflict of interest statement
The authors declare they have no conflict of interest.
Figures
References
-
- Stommel JM, Kimmelman AC, Ying H, Nabioullin R, Ponugoti AH, Wiedemeyer R, Stegh AH, Bradner JE, Ligon KL, Brennan C, Chin L, DePinho RA. Coactivation of receptor tyrosine kinases affects the response of tumor cells to targeted therapies. Science. 2007;318:287–90. - PubMed
-
- Snuderl M, Fazlollahi L, Le LP, Nitta M, Zhelyazkova BH, Davidson CJ, Akhavanfard S, Cahill DP, Aldape KD, Betensky RA, Louis DN, Iafrate AJ. Mosaic amplification of multiple receptor tyrosine kinase genes in glioblastoma. Cancer Cell. 2011;20:810–7. - PubMed
-
- Szerlip NJ, Pedraza A, Chakravarty D, Azim M, McGuire J, Fang Y, Ozawa T, Holland EC, Huse JT, Jhanwar S, Leversha MA, Mikkelsen T, Brennan CW. Intratumoral heterogeneity of receptor tyrosine kinases EGFR and PDGFRA amplification in glioblastoma defines subpopulations with distinct growth factor response. Proc Natl Acad Sci U S A. 2012;109:3041–6. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
