

Sortilin and SorLA display distinct roles in processing and trafficking of amyloid precursor protein
- PMID: 23283322
- PMCID: PMC6618638
- DOI: 10.1523/JNEUROSCI.2371-12.2013
Sortilin and SorLA display distinct roles in processing and trafficking of amyloid precursor protein
Abstract
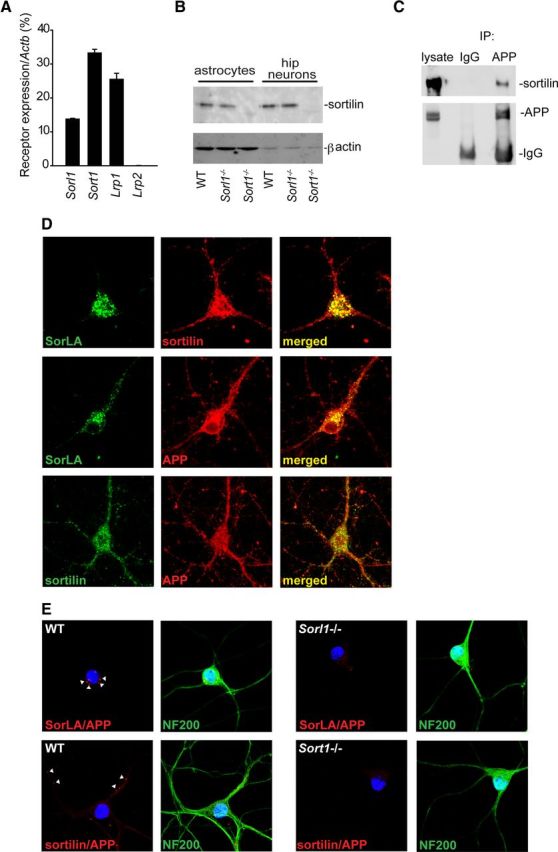
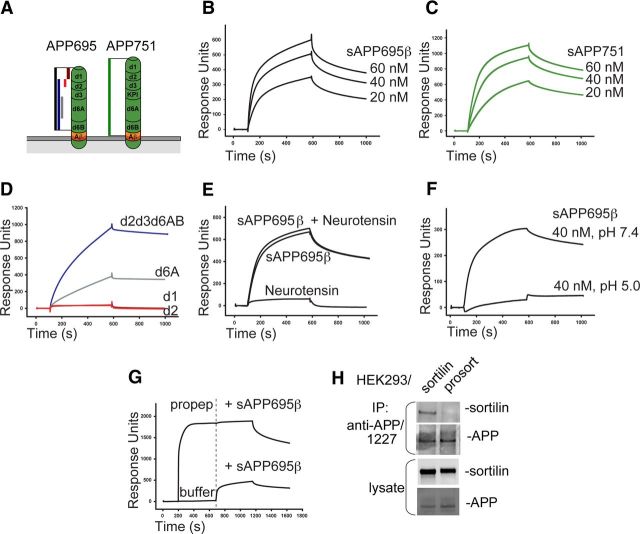
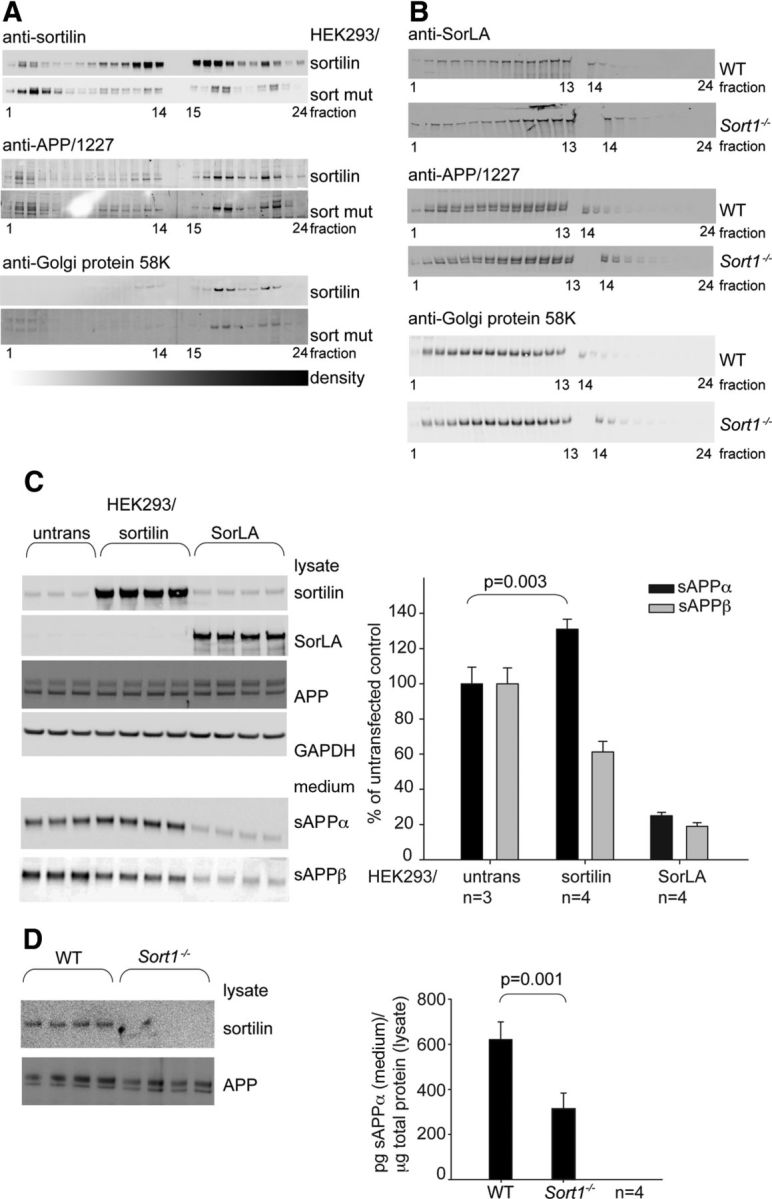
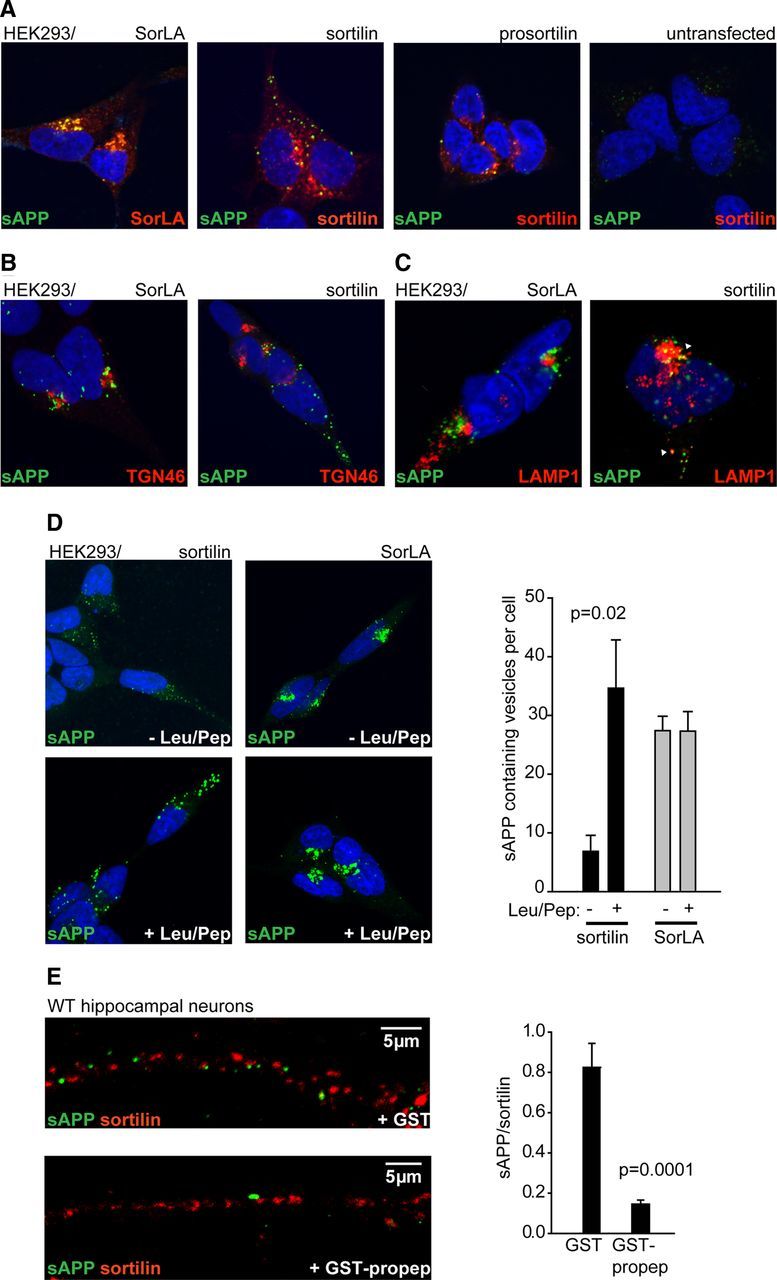
The development and progression of Alzheimer's disease is linked to excessive production of toxic amyloid-β peptide, initiated by β-secretase cleavage of the amyloid precursor protein (APP). In contrast, soluble APPα (sAPPα) generated by the α-secretase is known to stimulate dendritic branching and enhance synaptic function. Regulation of APP processing, and the shift from neurotrophic to neurotoxic APP metabolism remains poorly understood, but the cellular localization of APP and its interaction with various receptors is considered important. We here identify sortilin as a novel APP interaction partner. Like the related APP receptor SorLA, sortilin is highly expressed in the CNS, but whereas SorLA mainly colocalizes with APP in the soma, sortilin interacts with APP in neurites. The presence of sortilin promotes α-secretase cleavage of APP, unlike SorLA, which inhibits the generation of all soluble products. Also, sortilin and SorLA both bind and mediate internalization of sAPP but to different cellular compartments. The interaction involves the 6A domain of APP, present in both neuronal and non-neuronal APP isoforms. This is important as sAPP receptors described so far only bind the non-neuronal isoforms, leaving SorLA and sortilin as the only receptors for sAPP generated by neurons. Together, our findings establish sortilin, as a novel APP interaction partner that influences both production and cellular uptake of sAPP.
Figures
References
-
- Andersen OM, Reiche J, Schmidt V, Gotthardt M, Spoelgen R, Behlke J, von Arnim CA, Breiderhoff T, Jansen P, Wu X, Bales KR, Cappai R, Masters CL, Gliemann J, Mufson EJ, Hyman BT, Paul SM, Nykjaer A, Willnow TE. Neuronal sorting protein-related receptor sorLA/LR11 regulates processing of the amyloid precursor protein. Proc Natl Acad Sci U S A. 2005;102:13461–13466. - PMC - PubMed
-
- Andersen OM, Schmidt V, Spoelgen R, Gliemann J, Behlke J, Galatis D, McKinstry WJ, Parker MW, Masters CL, Hyman BT, Cappai R, Willnow TE. Molecular dissection of the interaction between amyloid precursor protein and its neuronal trafficking receptor SorLA/LR11. Biochemistry. 2006;45:2618–2628. - PubMed
-
- Bour A, Little S, Dodart JC, Kelche C, Mathis C. A secreted form of the beta-amyloid precursor protein (sAPP695) improves spatial recognition memory in OF1 mice. Neurobiol Learn Mem. 2004;81:27–38. - PubMed
-
- Cam JA, Zerbinatti CV, Knisely JM, Hecimovic S, Li Y, Bu G. The low density lipoprotein receptor-related protein 1B retains beta-amyloid precursor protein at the cell surface and reduces amyloid-beta peptide production. J Biol Chem. 2004;279:29639–29646. - PubMed
-
- Cramer JF, Gustafsen C, Behrens MA, Oliveira CL, Pedersen JS, Madsen P, Petersen CM, Thirup SS. GGA autoinhibition revisited. Traffic. 2010;11:259–273. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases