

Testing the sorption hypothesis in olfaction: a limited role for sniff strength in shaping primary odor representations during behavior
- PMID: 23283324
- PMCID: PMC3710131
- DOI: 10.1523/JNEUROSCI.4101-12.2013
Testing the sorption hypothesis in olfaction: a limited role for sniff strength in shaping primary odor representations during behavior
Abstract
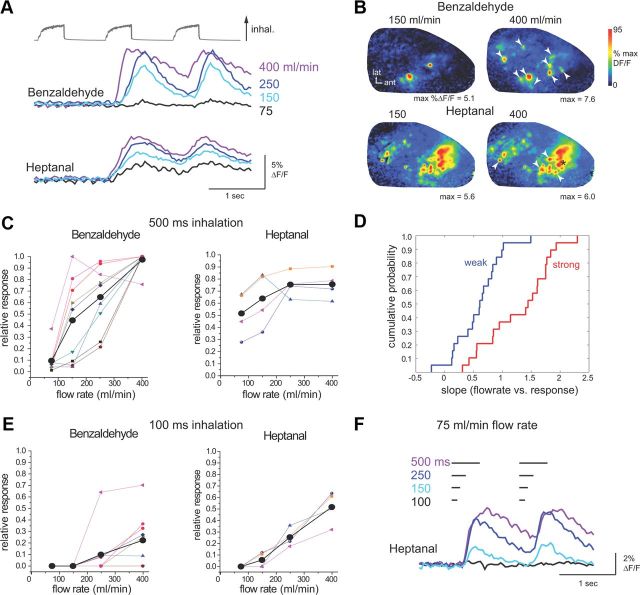
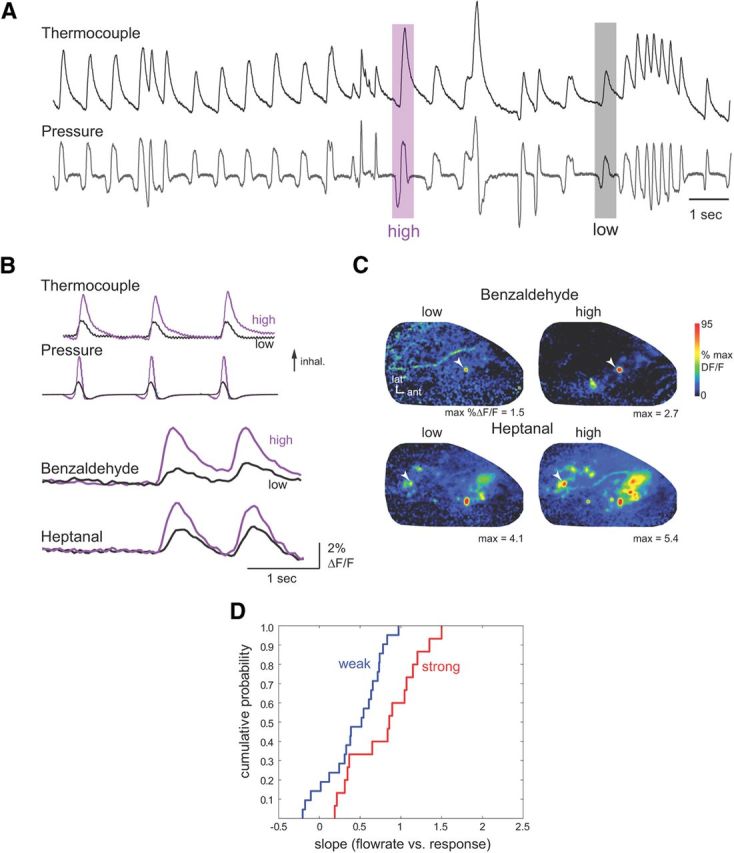
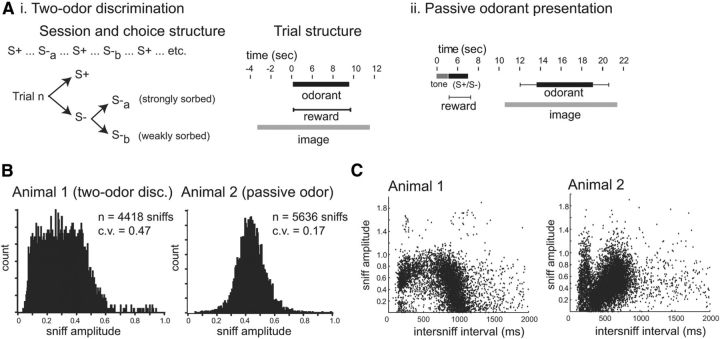
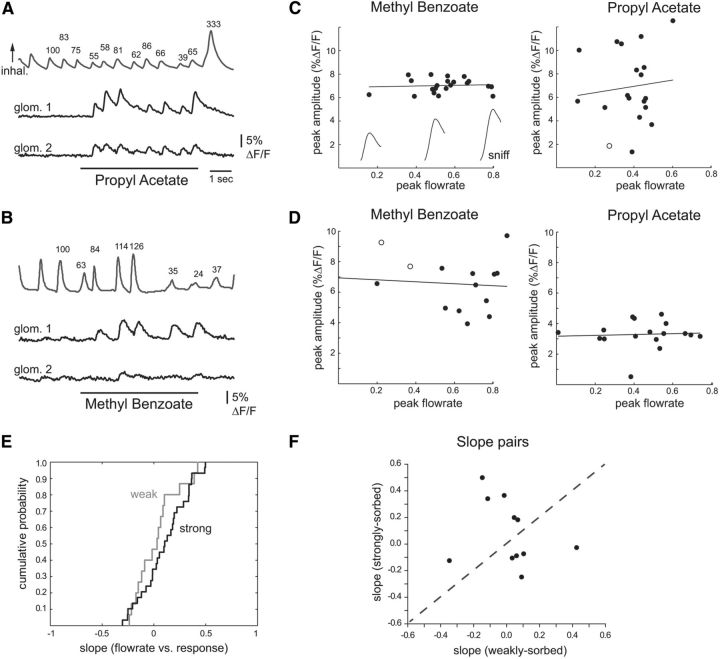
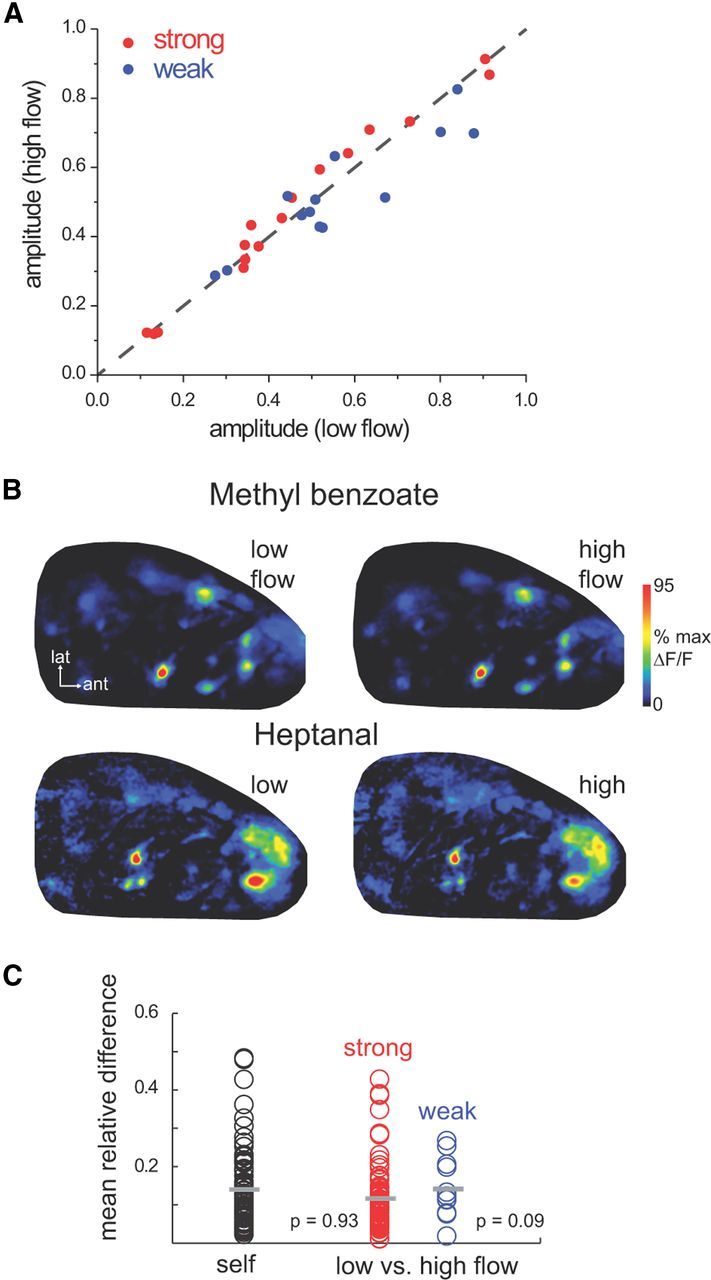
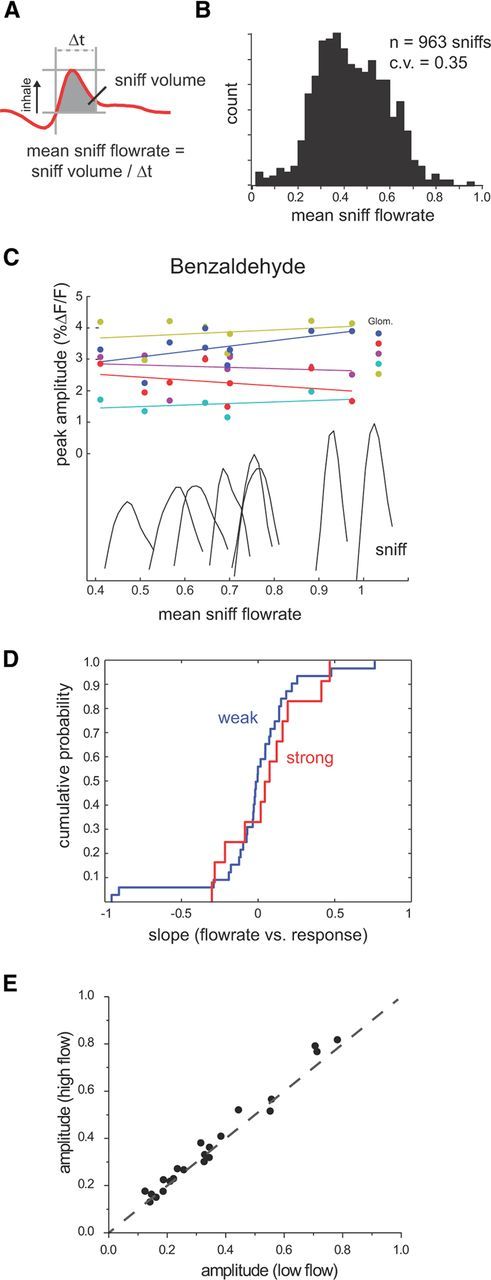
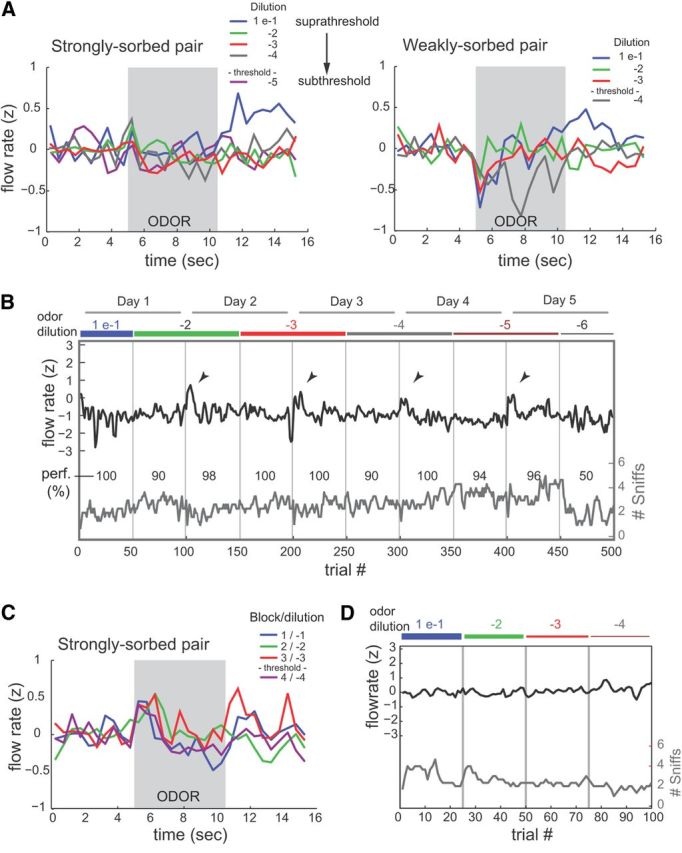
The acquisition of sensory information during behavior shapes the neural representation, central processing, and perception of external stimuli. In mammals, a sniff represents the basic unit of odor sampling, yet how sniffing shapes odor representations remains poorly understood. Perhaps the earliest hypothesis of the role of sniffing in olfaction arises from the fact that odorants with different physicochemical properties exhibit different patterns of deposition across the olfactory epithelium, and that these patterns are differentially affected by flow rate. However, whether sniff flow rates shape odor representations during natural sniffing remains untested, and whether animals make use of odorant sorption-airflow relationships as part of an active odor-sampling strategy remains unclear. We tested these ideas in the intact rat using a threefold approach. First, we asked whether sniff strength shapes odor representations in vivo by imaging from olfactory receptor neuron (ORN) terminals during controlled changes in inhalation flow in the anesthetized rat. Second, we asked whether sniff strength shapes odor representations by imaging from ORNs during natural sniffing in the awake rat. Third, we asked whether rats actively modulate sniff strength during an odor discrimination task. We found that, while artificial changes in flow rate can alter ORN responses, sniff strength has negligible effect on odor representations during natural sniffing, and behaving rats do not modulate flow rate to improve odor discrimination. These data suggest that modulating sniff strength does not shape odor representations sufficiently to be part of a strategy for active odor sensing in the behaving animal.
Figures
References
-
- Adrian ED. Sensory discrimination with some recent evidence from the olfactory organ. Br Med Bull. 1950;6:330–333. - PubMed
-
- Amoore J, Buttery R. Partition coefficients and comparative olfactometry. Chem Senses and Flavour. 1978;3:57–71.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources