

Genetic analysis of synaptotagmin C2 domain specificity in regulating spontaneous and evoked neurotransmitter release
- PMID: 23283333
- PMCID: PMC3711629
- DOI: 10.1523/JNEUROSCI.3214-12.2013
Genetic analysis of synaptotagmin C2 domain specificity in regulating spontaneous and evoked neurotransmitter release
Abstract
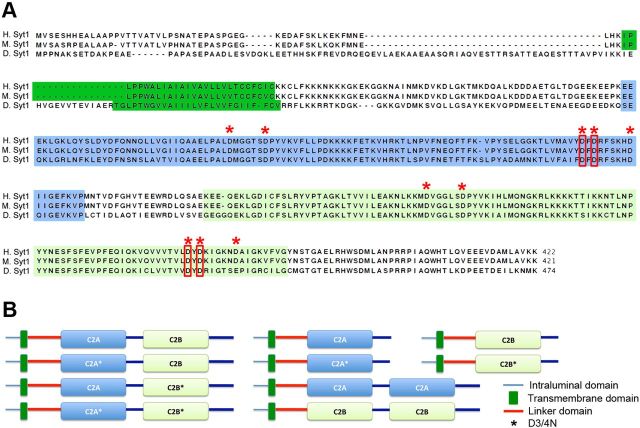
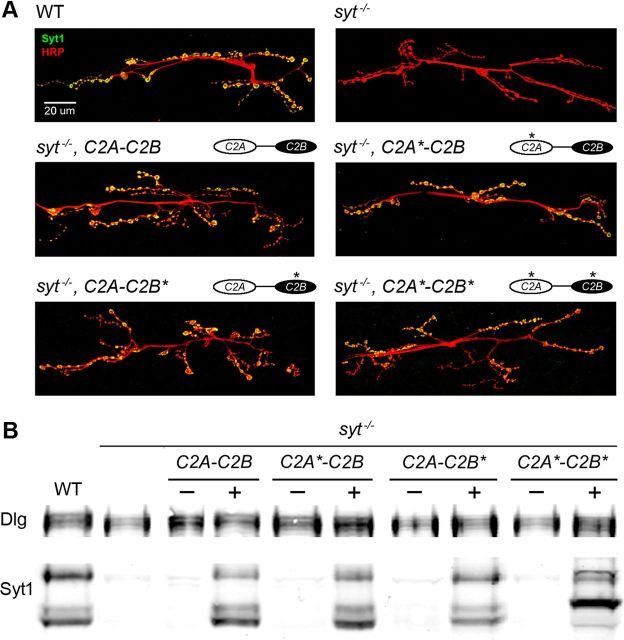
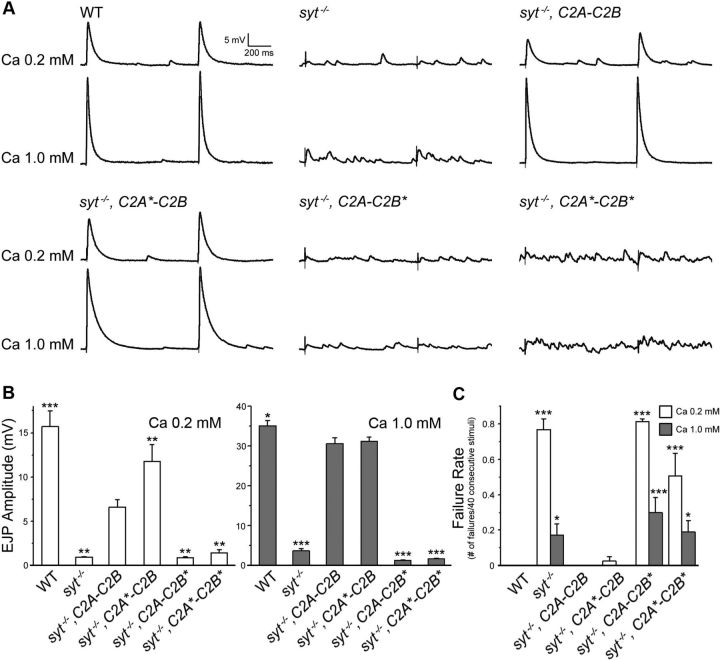
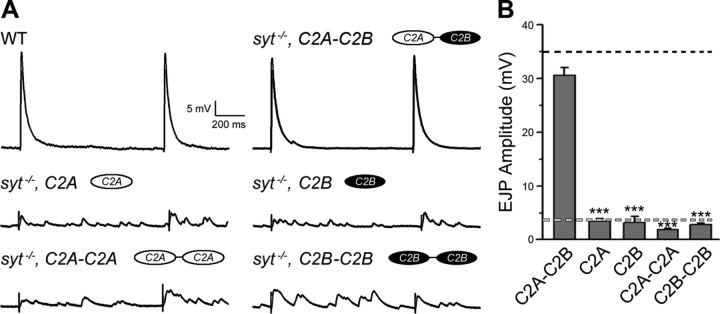
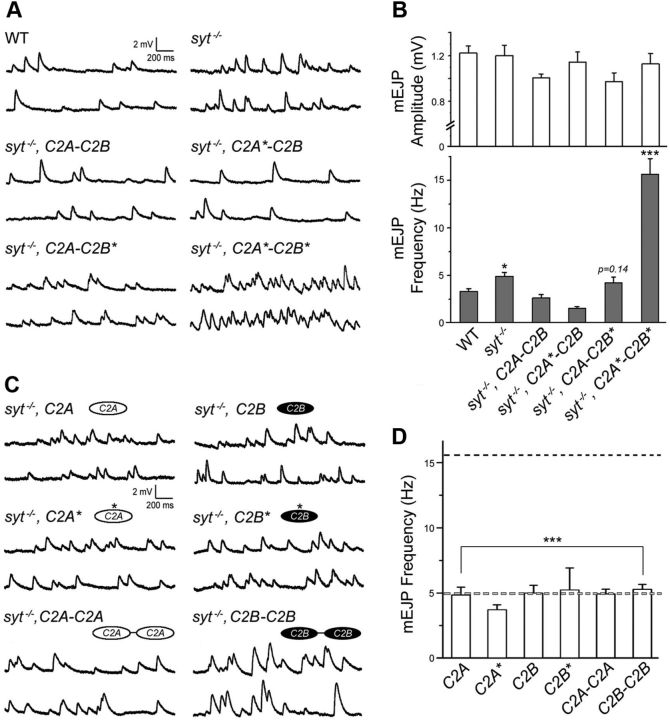
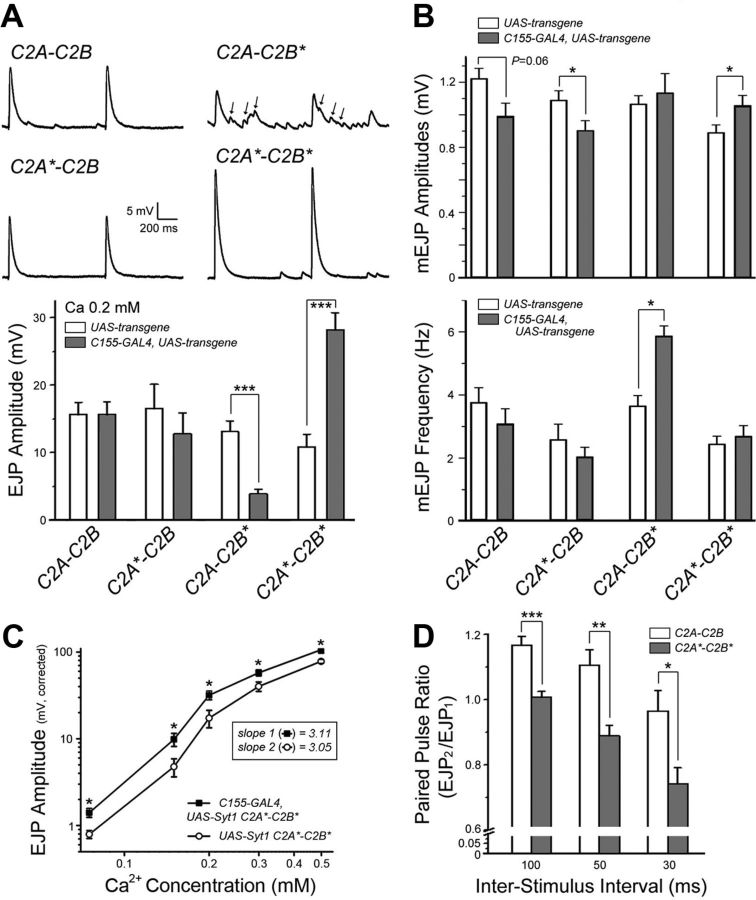
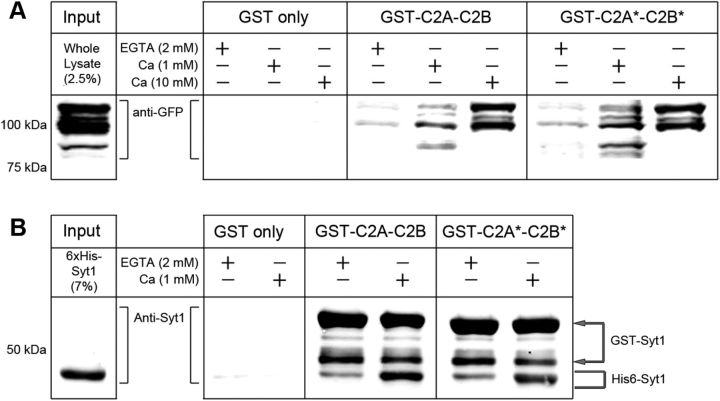
Synaptic vesicle fusion mediates communication between neurons and is triggered by rapid influx of Ca(2+). The Ca(2+)-triggering step for fusion is regulated by the synaptic vesicle transmembrane protein Synaptotagmin 1 (Syt1). Syt1 contains two cytoplasmic C2 domains, termed C2A and C2B, which coordinate Ca(2+) binding. Although C2A and C2B share similar topology, binding of Ca(2+) ions to the C2B domain has been suggested as the only critical trigger for evoked vesicle release. If and how C2A domain function is coordinated with C2B remain unclear. In this study, we generated a panel of Syt1 chimeric constructs in Drosophila to delineate the unique and shared functions of each C2 domain in regulation of synaptic vesicle fusion. Expression of Syt 1 transgenes containing only individual C2 domains, or dual C2A-C2A or C2B-C2B chimeras, failed to restore Syt1 function in a syt1(-/-) null mutant background, indicating both C2A and C2B are specifically required to support fast synchronous release. Mutations that disrupted Ca(2+) binding to both C2 domains failed to rescue evoked release, but supported synaptic vesicle docking and endocytosis, indicating that these functions of Syt1 are Ca(2+)-independent. The dual C2 domain Ca(2+)-binding mutant also enhanced spontaneous fusion while dramatically increasing evoked release when coexpressed with native Syt1. Together, these data indicate that synaptic transmission can be regulated by Syt1 multimerization and that both C2 domains of Syt1 are uniquely required for modulating Ca(2+)-independent spontaneous fusion and Ca(2+)-dependent synchronous release.
Figures


References
-
- Bai J, Earles CA, Lewis JL, Chapman ER. Membrane-embedded synaptotagmin penetrates cis or trans target membranes and clusters via a novel mechanism. J Biol Chem. 2000;275:25427–25435. - PubMed
-
- Bai J, Wang CT, Richards DA, Jackson MB, Chapman ER. Fusion pore dynamics are regulated by synaptotagmin*t-SNARE interactions. Neuron. 2004;41:929–942. - PubMed
-
- Bennett MK, Calakos N, Scheller RH. Syntaxin: a synaptic protein implicated in docking of synaptic vesicles at presynaptic active zones. Science. 1992;257:255–259. - PubMed
-
- Brand AH, Perrimon N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development. 1993;118:401–415. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous