

Prototype foamy virus protease activity is essential for intraparticle reverse transcription initiation but not absolutely required for uncoating upon host cell entry
- PMID: 23283957
- PMCID: PMC3592149
- DOI: 10.1128/JVI.02323-12
Prototype foamy virus protease activity is essential for intraparticle reverse transcription initiation but not absolutely required for uncoating upon host cell entry
Abstract
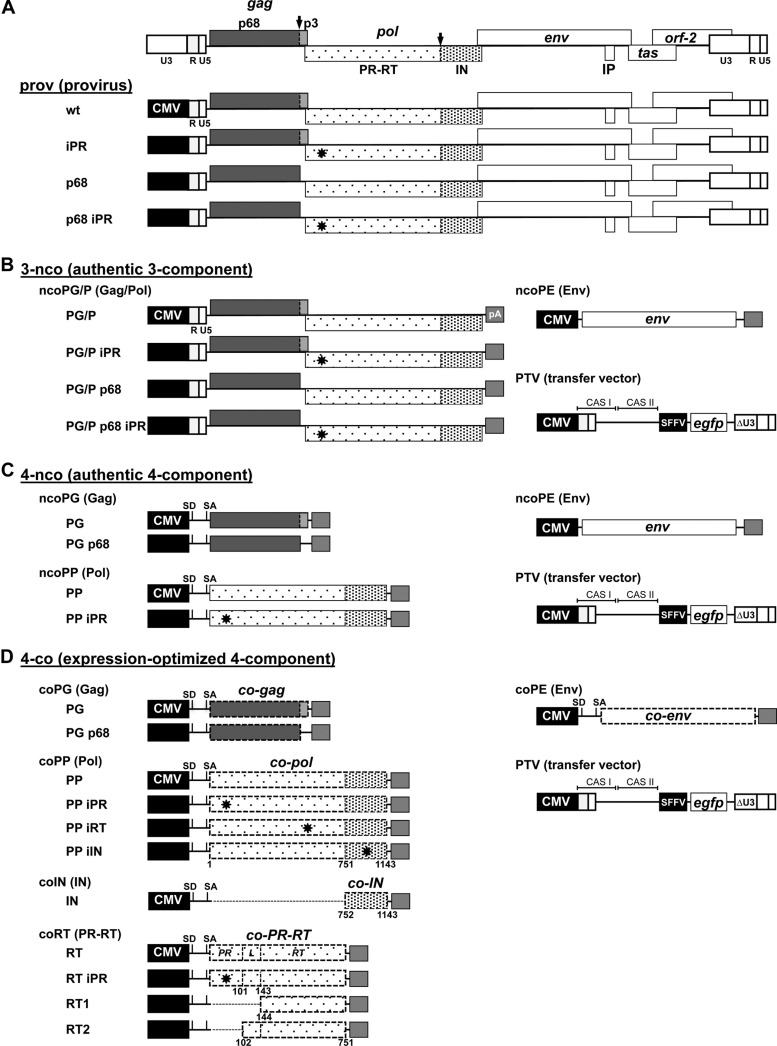
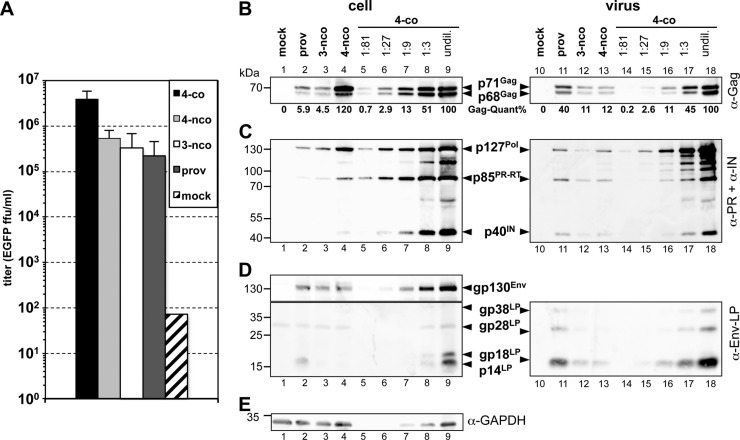
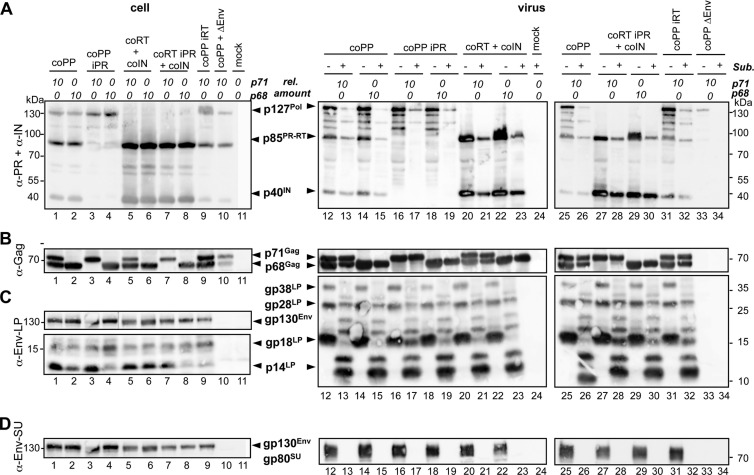
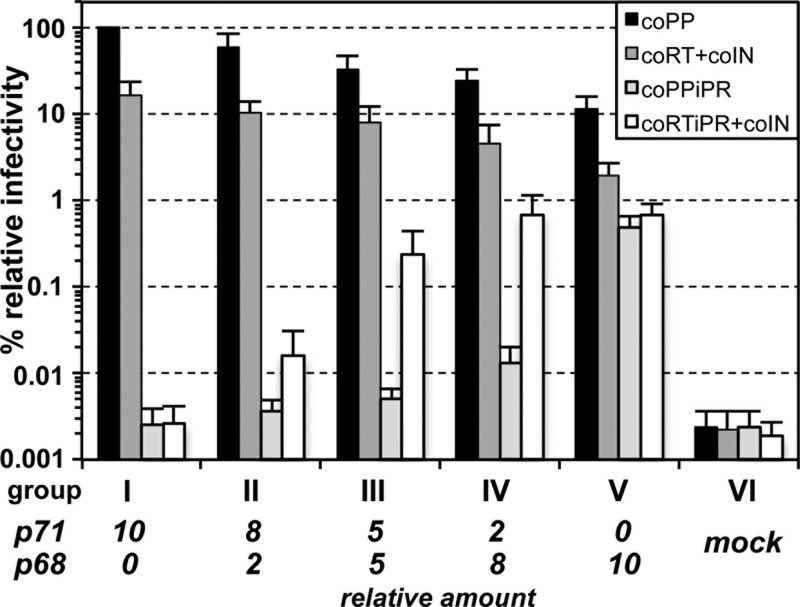
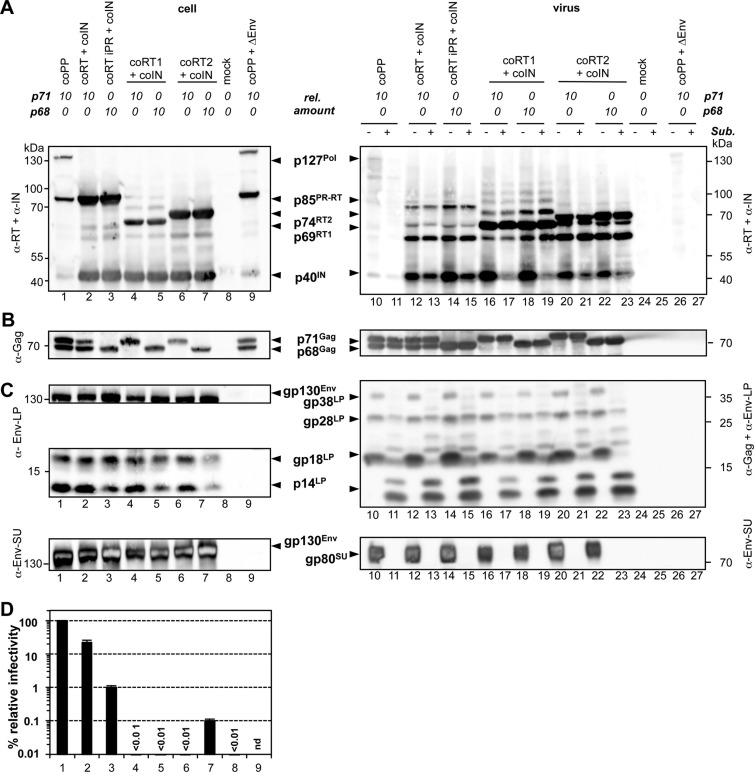
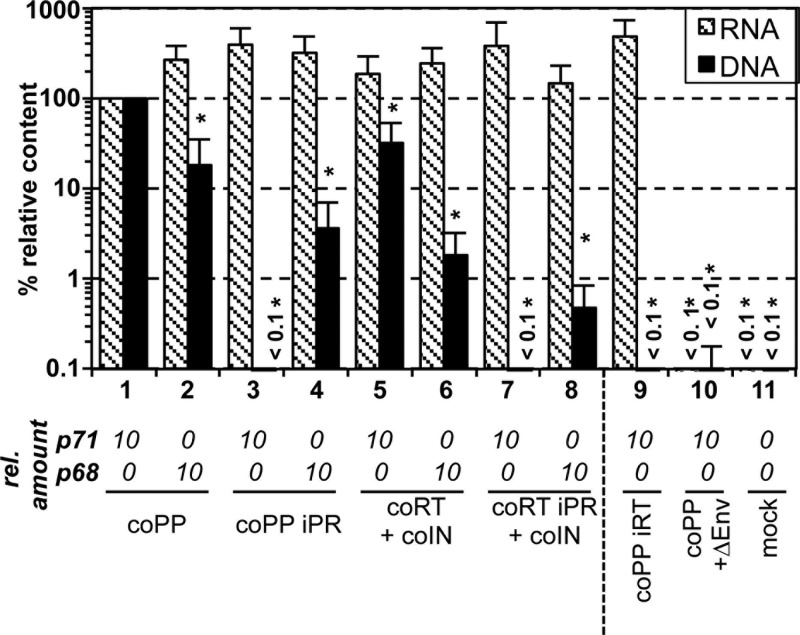
Foamy viruses (FVs) are unique among retroviruses in performing genome reverse transcription (RTr) late in replication, resulting in an infectious DNA genome, and also in their unusual Pol biosynthesis and encapsidation strategy. In addition, FVs display only very limited Gag and Pol processing by the viral protease (PR) during particle morphogenesis and disassembly, both thought to be crucial for viral infectivity. Here, we report the generation of functional prototype FV (PFV) particles from mature or partially processed viral capsid and enzymatic proteins with infectivity levels of up to 20% of the wild type. Analysis of protein and nucleic acid composition, as well as infectivity, of virions generated from different Gag and Pol combinations (including both expression-optimized and authentic PFV open reading frames [ORFs]) revealed that precursor processing of Gag, but not Pol, during particle assembly is essential for production of infectious virions. Surprisingly, when processed Gag (instead of Gag precursor) was provided together with PR-deficient Pol precursor during virus production, infectious, viral DNA-containing particles were obtained, even when different vector or proviral expression systems were used. Although virion infectivity was reduced to 0.5 to 2% relative to that of the respective parental constructs, this finding overturns the current dogma in the FV literature that viral PR activity is absolutely essential at some point during target cell entry. Furthermore, it demonstrates that viral PR-mediated Gag precursor processing during particle assembly initiates intraparticle RTr. Finally, it shows that reverse transcriptase (RT) and integrase are enzymatically active in the Pol precursor within the viral capsid, thus enabling productive host cell infection.
Figures


Similar articles
-
Orthoretroviral-like prototype foamy virus Gag-Pol expression is compatible with viral replication.Retrovirology. 2011 Aug 15;8:66. doi: 10.1186/1742-4690-8-66. Retrovirology. 2011. PMID: 21843316 Free PMC article.
-
The cooperative function of arginine residues in the Prototype Foamy Virus Gag C-terminus mediates viral and cellular RNA encapsidation.Retrovirology. 2014 Oct 8;11:87. doi: 10.1186/s12977-014-0087-7. Retrovirology. 2014. PMID: 25292281 Free PMC article.
-
Novel functions of prototype foamy virus Gag glycine- arginine-rich boxes in reverse transcription and particle morphogenesis.J Virol. 2011 Feb;85(4):1452-63. doi: 10.1128/JVI.01731-10. Epub 2010 Nov 24. J Virol. 2011. PMID: 21106749 Free PMC article.
-
Foamy virus assembly with emphasis on pol encapsidation.Viruses. 2013 Mar 20;5(3):886-900. doi: 10.3390/v5030886. Viruses. 2013. PMID: 23518575 Free PMC article. Review.
-
Proteolytic processing of foamy virus Gag and Pol proteins.Curr Top Microbiol Immunol. 2003;277:63-88. doi: 10.1007/978-3-642-55701-9_3. Curr Top Microbiol Immunol. 2003. PMID: 12908768 Review.
Cited by
-
Structural and Functional Aspects of Foamy Virus Protease-Reverse Transcriptase.Viruses. 2019 Jul 2;11(7):598. doi: 10.3390/v11070598. Viruses. 2019. PMID: 31269675 Free PMC article. Review.
-
Functional Analyses of Bovine Foamy Virus-Encoded miRNAs Reveal the Importance of a Defined miRNA for Virus Replication and Host-Virus Interaction.Viruses. 2020 Nov 2;12(11):1250. doi: 10.3390/v12111250. Viruses. 2020. PMID: 33147813 Free PMC article.
-
Structure of a Spumaretrovirus Gag Central Domain Reveals an Ancient Retroviral Capsid.PLoS Pathog. 2016 Nov 9;12(11):e1005981. doi: 10.1371/journal.ppat.1005981. eCollection 2016 Nov. PLoS Pathog. 2016. PMID: 27829070 Free PMC article.
-
Foamy Virus Protein-Nucleic Acid Interactions during Particle Morphogenesis.Viruses. 2016 Aug 30;8(9):243. doi: 10.3390/v8090243. Viruses. 2016. PMID: 27589786 Free PMC article. Review.
-
Crystal Structure of a Retroviral Polyprotein: Prototype Foamy Virus Protease-Reverse Transcriptase (PR-RT).Viruses. 2021 Jul 29;13(8):1495. doi: 10.3390/v13081495. Viruses. 2021. PMID: 34452360 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
