

Visualization of the exocyst complex dynamics at the plasma membrane of Arabidopsis thaliana
- PMID: 23283982
- PMCID: PMC3571873
- DOI: 10.1091/mbc.E12-06-0492
Visualization of the exocyst complex dynamics at the plasma membrane of Arabidopsis thaliana
Abstract
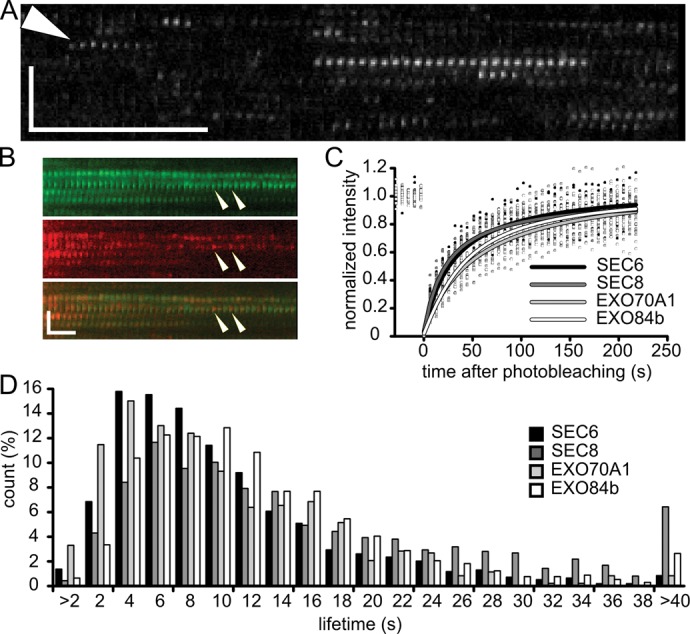
The exocyst complex, an effector of Rho and Rab GTPases, is believed to function as an exocytotic vesicle tether at the plasma membrane before soluble N-ethylmaleimide-sensitive factor attachment protein receptor (SNARE) complex formation. Exocyst subunits localize to secretory-active regions of the plasma membrane, exemplified by the outer domain of Arabidopsis root epidermal cells. Using variable-angle epifluorescence microscopy, we visualized the dynamics of exocyst subunits at this domain. The subunits colocalized in defined foci at the plasma membrane, distinct from endocytic sites. Exocyst foci were independent of cytoskeleton, although prolonged actin disruption led to changes in exocyst localization. Exocyst foci partially overlapped with vesicles visualized by VAMP721 v-SNARE, but the majority of the foci represent sites without vesicles, as indicated by electron microscopy and drug treatments, supporting the concept of the exocyst functioning as a dynamic particle. We observed a decrease of SEC6-green fluorescent protein foci in an exo70A1 exocyst mutant. Finally, we documented decreased VAMP721 trafficking to the plasma membrane in exo70A1 and exo84b mutants. Our data support the concept that the exocyst-complex subunits dynamically dock and undock at the plasma membrane to create sites primed for vesicle tethering.
Figures





Similar articles
-
Synergy among Exocyst and SNARE Interactions Identifies a Functional Hierarchy in Secretion during Vegetative Growth.Plant Cell. 2020 Sep;32(9):2951-2963. doi: 10.1105/tpc.20.00280. Epub 2020 Jul 22. Plant Cell. 2020. PMID: 32699172 Free PMC article.
-
The exocyst complex contributes to PIN auxin efflux carrier recycling and polar auxin transport in Arabidopsis.Plant J. 2013 Mar;73(5):709-19. doi: 10.1111/tpj.12074. Epub 2013 Jan 10. Plant J. 2013. PMID: 23163883
-
The small GTPase RABA2a recruits SNARE proteins to regulate the secretory pathway in parallel with the exocyst complex in Arabidopsis.Mol Plant. 2022 Mar 7;15(3):398-418. doi: 10.1016/j.molp.2021.11.008. Epub 2021 Nov 16. Mol Plant. 2022. PMID: 34798312
-
Regulation of Cell Polarity by Exocyst-Mediated Trafficking.Cold Spring Harb Perspect Biol. 2018 Mar 1;10(3):a031401. doi: 10.1101/cshperspect.a031401. Cold Spring Harb Perspect Biol. 2018. PMID: 28264817 Free PMC article. Review.
-
Exposing the Elusive Exocyst Structure.Trends Biochem Sci. 2018 Sep;43(9):714-725. doi: 10.1016/j.tibs.2018.06.012. Epub 2018 Jul 25. Trends Biochem Sci. 2018. PMID: 30055895 Free PMC article. Review.
Cited by
-
Exocyst Subunit EXO70H4 Has a Specific Role in Callose Synthase Secretion and Silica Accumulation.Plant Physiol. 2018 Mar;176(3):2040-2051. doi: 10.1104/pp.17.01693. Epub 2018 Jan 4. Plant Physiol. 2018. PMID: 29301954 Free PMC article.
-
CSI1, PATROL1, and exocyst complex cooperate in delivery of cellulose synthase complexes to the plasma membrane.Proc Natl Acad Sci U S A. 2018 Apr 10;115(15):E3578-E3587. doi: 10.1073/pnas.1800182115. Epub 2018 Mar 26. Proc Natl Acad Sci U S A. 2018. PMID: 29581258 Free PMC article.
-
Interplay of EXO70 and MLO proteins modulates trichome cell wall composition and susceptibility to powdery mildew.Plant Cell. 2024 Mar 29;36(4):1007-1035. doi: 10.1093/plcell/koad319. Plant Cell. 2024. PMID: 38124479 Free PMC article.
-
The exocyst at the interface between cytoskeleton and membranes in eukaryotic cells.Front Plant Sci. 2014 Jan 2;4:543. doi: 10.3389/fpls.2013.00543. Front Plant Sci. 2014. PMID: 24427163 Free PMC article. Review.
-
Endosidin2 targets conserved exocyst complex subunit EXO70 to inhibit exocytosis.Proc Natl Acad Sci U S A. 2016 Jan 5;113(1):E41-50. doi: 10.1073/pnas.1521248112. Epub 2015 Nov 25. Proc Natl Acad Sci U S A. 2016. PMID: 26607451 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
