

High-pressure NMR reveals close similarity between cold and alcohol protein denaturation in ubiquitin
- PMID: 23284170
- PMCID: PMC3562818
- DOI: 10.1073/pnas.1212222110
High-pressure NMR reveals close similarity between cold and alcohol protein denaturation in ubiquitin
Abstract
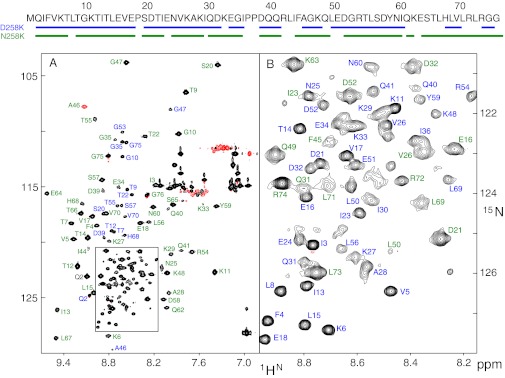
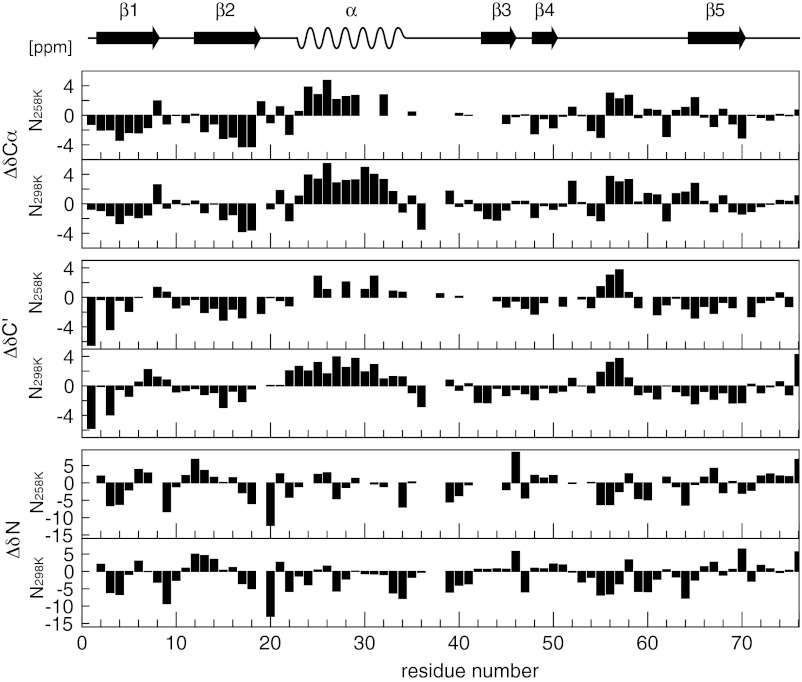
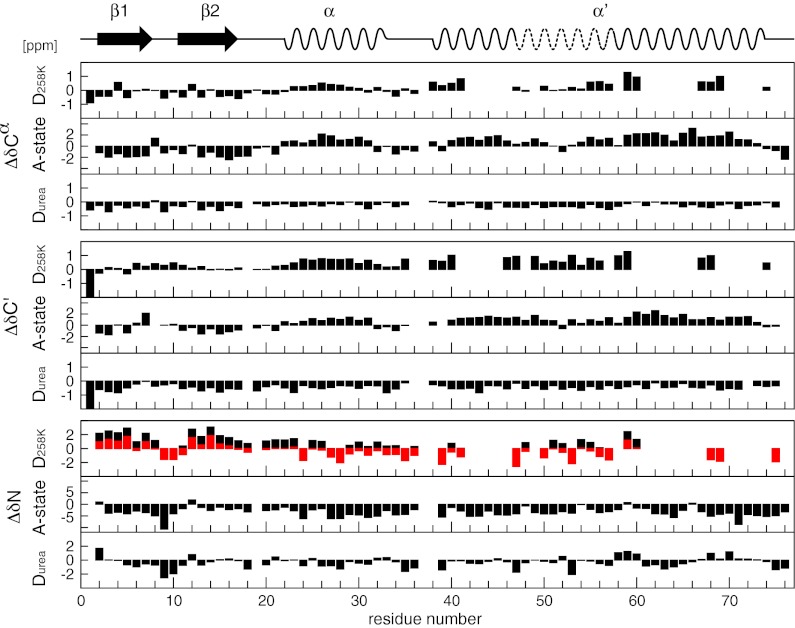
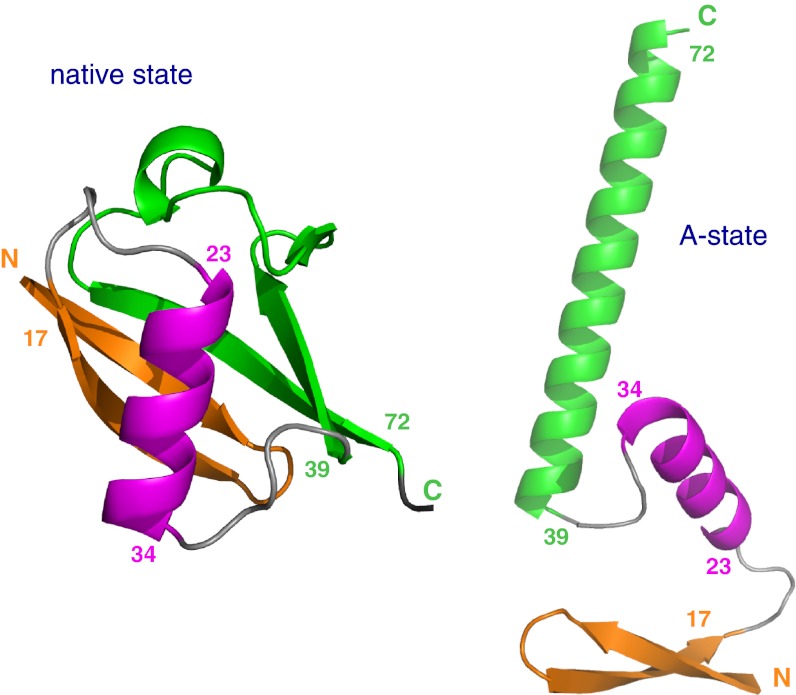
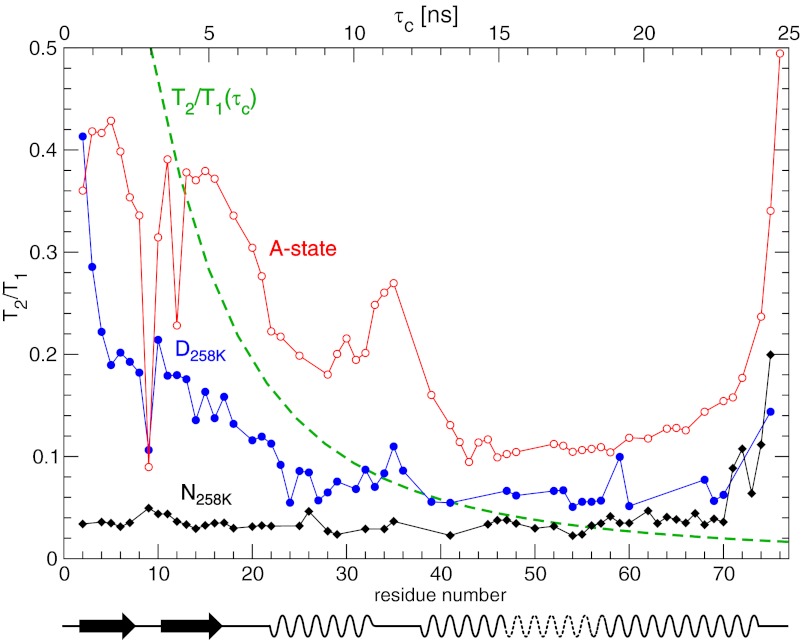
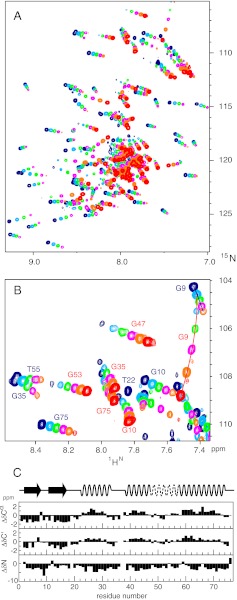
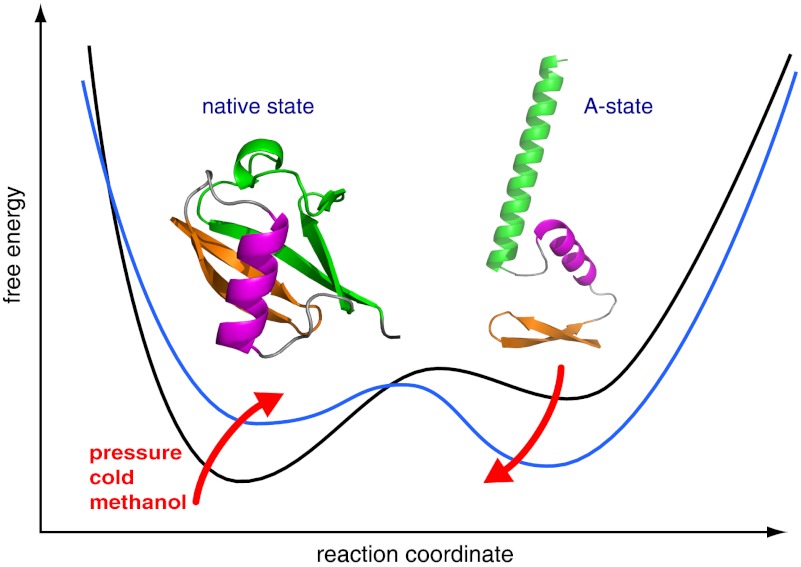
Proteins denature not only at high, but also at low temperature as well as high pressure. These denatured states are not easily accessible for experiment, because usually heat denaturation causes aggregation, whereas cold or pressure denaturation occurs at temperatures well below the freezing point of water or pressures above 5 kbar, respectively. Here we have obtained atomic details of the pressure-assisted, cold-denatured state of ubiquitin at 2,500 bar and 258 K by high-resolution NMR techniques. Under these conditions, a folded, native-like and a disordered state exist in slow exchange. Secondary chemical shifts show that the disordered state has structural propensities for a native-like N-terminal β-hairpin and α-helix and a nonnative C-terminal α-helix. These propensities are very similar to the previously described alcohol-denatured (A-)state. Similar to the A-state, (15)N relaxation data indicate that the secondary structure elements move as independent segments. The close similarity of pressure-assisted, cold-denatured, and alcohol-denatured states with native and nonnative secondary elements supports a hierarchical mechanism of folding and supports the notion that similar to alcohol, pressure and cold reduce the hydrophobic effect. Indeed, at nondenaturing concentrations of methanol, a complete transition from the native to the A-state can be achieved at ambient temperature by varying the pressure from 1 to 2,500 bar. The methanol-assisted pressure transition is completely reversible and can also be induced in protein G. This method should allow highly detailed studies of protein-folding transitions in a continuous and reversible manner.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Hawley SA. Reversible pressure—temperature denaturation of chymotrypsinogen. Biochemistry. 1971;10(13):2436–2442. - PubMed
-
- Privalov PL, Griko YuV, Venyaminov SYu, Kutyshenko VP. Cold denaturation of myoglobin. J Mol Biol. 1986;190(3):487–498. - PubMed
-
- Smeller L. Pressure-temperature phase diagrams of biomolecules. Biochim Biophys Acta. 2002;1595(1–2):11–29. - PubMed
-
- Kauzmann W. Some factors in the interpretation of protein denaturation. Adv Protein Chem. 1959;14:1–63. - PubMed
-
- Tanford C. Contribution of hydrophobic interactions to the stability of the globular conformation of proteins. J Am Chem Soc. 1962;84:4240–4247.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
