

Lack of phenotypic and evolutionary cross-resistance against parasitoids and pathogens in Drosophila melanogaster
- PMID: 23285247
- PMCID: PMC3528725
- DOI: 10.1371/journal.pone.0053002
Lack of phenotypic and evolutionary cross-resistance against parasitoids and pathogens in Drosophila melanogaster
Abstract
Background: When organisms are attacked by multiple natural enemies, the evolution of a resistance mechanism to one natural enemy will be influenced by the degree of cross-resistance to another natural enemy. Cross-resistance can be positive, when a resistance mechanism against one natural enemy also offers resistance to another; or negative, in the form of a trade-off, when an increase in resistance against one natural enemy results in a decrease in resistance against another. Using Drosophila melanogaster, an important model system for the evolution of invertebrate immunity, we test for the existence of cross-resistance against parasites and pathogens, at both a phenotypic and evolutionary level.
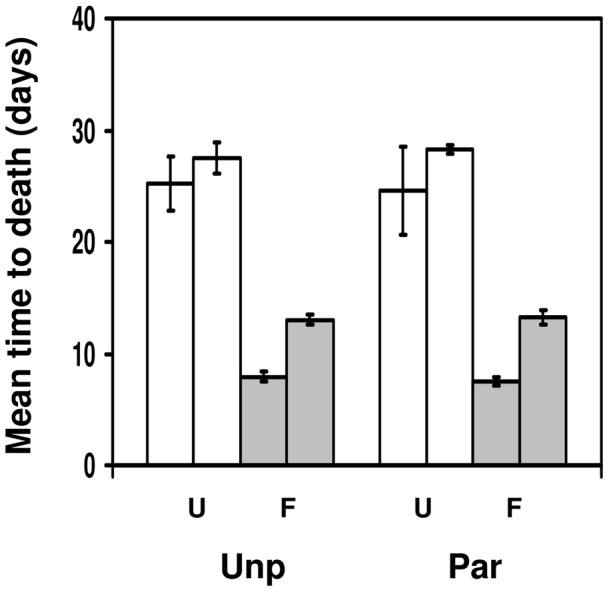
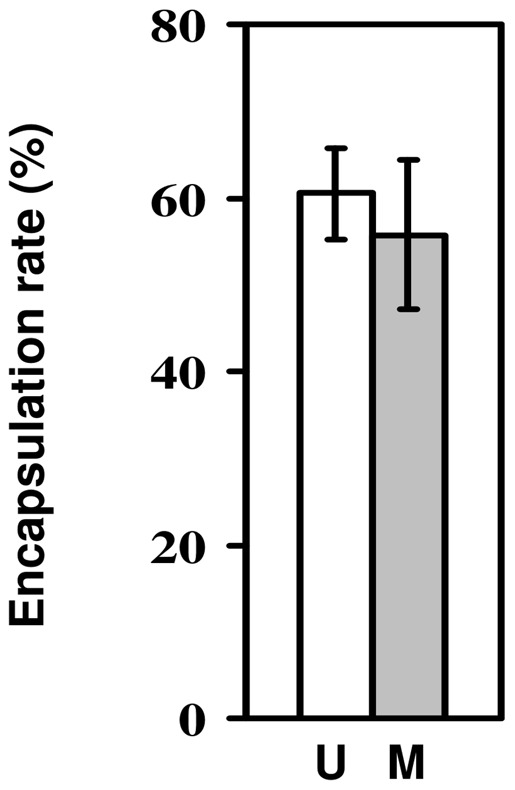
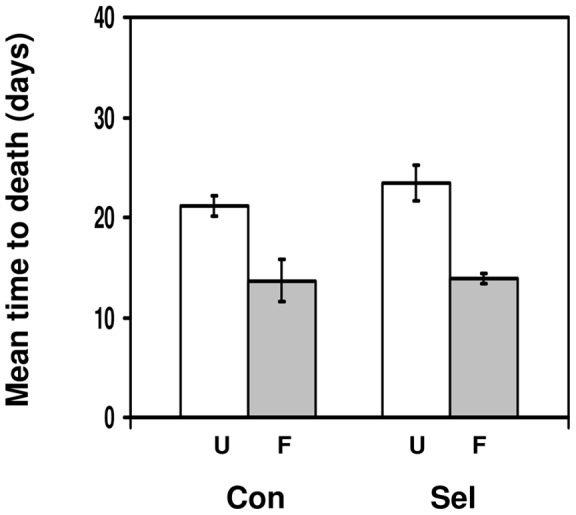
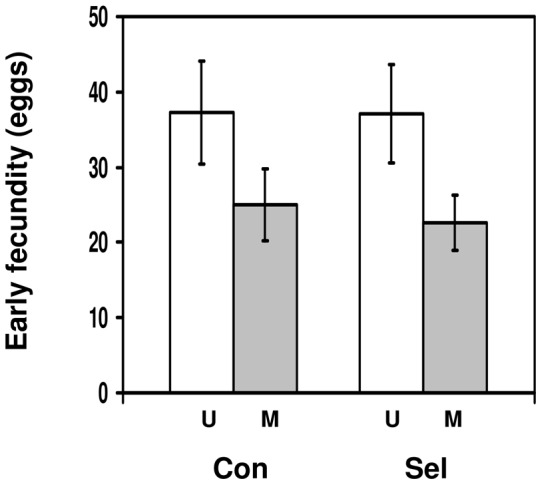
Methods: We used a field strain of D. melanogaster to test whether surviving parasitism by the parasitoid Asobara tabida has an effect on the resistance against Beauveria bassiana, an entomopathogenic fungus; and whether infection with the microsporidian Tubulinosema kingi has an effect on the resistance against A. tabida. We used lines selected for increased resistance to A. tabida to test whether increased parasitoid resistance has an effect on resistance against B. bassiana and T. kingi. We used lines selected for increased tolerance against B. bassiana to test whether increased fungal resistance has an effect on resistance against A. tabida.
Results/conclusions: We found no positive cross-resistance or trade-offs in the resistance to parasites and pathogens. This is an important finding, given the use of D. melanogaster as a model system for the evolution of invertebrate immunity. The lack of any cross-resistance to parasites and pathogens, at both the phenotypic and the evolutionary level, suggests that evolution of resistance against one class of natural enemies is largely independent of evolution of resistance against the other.
Conflict of interest statement
Figures
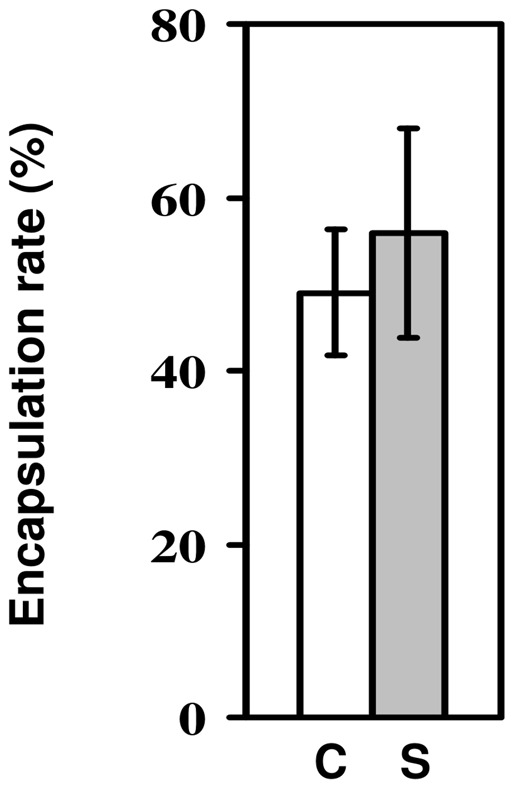
Similar articles
-
Experimental evolution shows Drosophila melanogaster resistance to a microsporidian pathogen has fitness costs.Evolution. 2009 Jan;63(1):104-14. doi: 10.1111/j.1558-5646.2008.00516.x. Epub 2008 Sep 11. Evolution. 2009. PMID: 18786186
-
CROSS-RESISTANCE FOLLOWING ARTIFICIAL SELECTION FOR INCREASED DEFENSE AGAINST PARASITOIDS IN DROSOPHILA MELANOGASTER.Evolution. 1999 Jun;53(3):966-972. doi: 10.1111/j.1558-5646.1999.tb05391.x. Evolution. 1999. PMID: 28565619
-
Evolutionary interactions between Drosophila melanogaster and its parasitoid Asobara tabida.Heredity (Edinb). 2000 Nov;85 Pt 5:450-8. doi: 10.1046/j.1365-2540.2000.00788.x. Heredity (Edinb). 2000. PMID: 11122423
-
Evolution of host resistance and parasitoid counter-resistance.Adv Parasitol. 2009;70:257-80. doi: 10.1016/S0065-308X(09)70010-7. Adv Parasitol. 2009. PMID: 19773074 Review.
-
Costs of resistance in insect-parasite and insect-parasitoid interactions.Parasitology. 2002;125 Suppl:S71-82. doi: 10.1017/s0031182002001750. Parasitology. 2002. PMID: 12622330 Review.
Cited by
-
Digenean-gastropod host associations inform on aspects of specific immunity in snails.Dev Comp Immunol. 2015 Feb;48(2):275-83. doi: 10.1016/j.dci.2014.06.014. Epub 2014 Jul 14. Dev Comp Immunol. 2015. PMID: 25034871 Free PMC article. Review.
-
Severe Insect Pest Impacts on New Zealand Pasture: The Plight of an Ecological Outlier.J Insect Sci. 2020 Mar 1;20(2):17. doi: 10.1093/jisesa/ieaa018. J Insect Sci. 2020. PMID: 32322881 Free PMC article. Review.
-
Meta-Analysis of Immune Induced Gene Expression Changes in Diverse Drosophila melanogaster Innate Immune Responses.Insects. 2022 May 23;13(5):490. doi: 10.3390/insects13050490. Insects. 2022. PMID: 35621824 Free PMC article.
-
Host adaptation is contingent upon the infection route taken by pathogens.PLoS Pathog. 2013;9(9):e1003601. doi: 10.1371/journal.ppat.1003601. Epub 2013 Sep 26. PLoS Pathog. 2013. PMID: 24086131 Free PMC article.
-
Coevolutionary dynamics of host-pathogen interaction with density-dependent mortality.J Math Biol. 2022 Jul 25;85(2):15. doi: 10.1007/s00285-022-01782-8. J Math Biol. 2022. PMID: 35877051 Free PMC article.
References
-
- Levin DA (1976) The chemical defenses of plants to pathogens and herbivores. Ann Rev Ecol Syst 7: 121–159.
-
- Maleck K, Dietrich PA (1999) Defense on multiple fronts: how do plants cope with diverse enemies? Trends Plant Sci 4: 215–219. - PubMed
-
- Hunter GW, Weinmann CJ, Hoffmann RG (2004) Studies on schistosomiasis. XVII. Non-reciprocal acquired resistance between Schistosoma mansoni and Schistosomatium douthitti in mice. Exp Parasitol 11: 133–140. - PubMed
-
- Kurtz J, Franz K (2003) Evidence for memory in invertebrate immunity. Nature 425: 37–38. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
