

Characterizing the importance of the biotin carboxylase domain dimer for Staphylococcus aureus pyruvate carboxylase catalysis
- PMID: 23286247
- PMCID: PMC3553216
- DOI: 10.1021/bi301294d
Characterizing the importance of the biotin carboxylase domain dimer for Staphylococcus aureus pyruvate carboxylase catalysis
Abstract
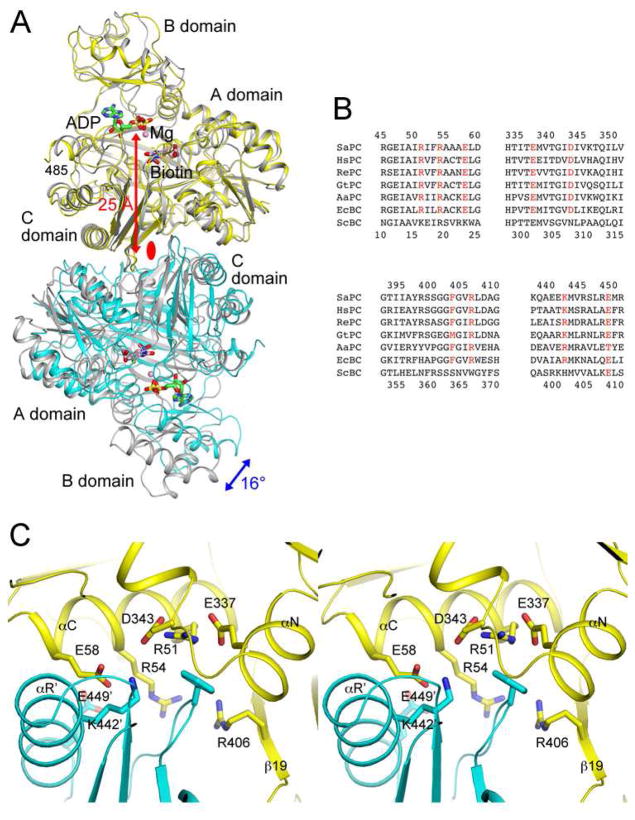
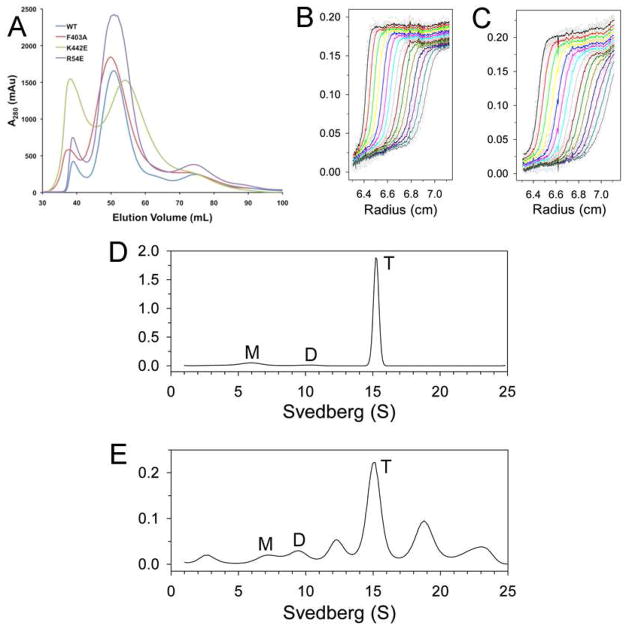
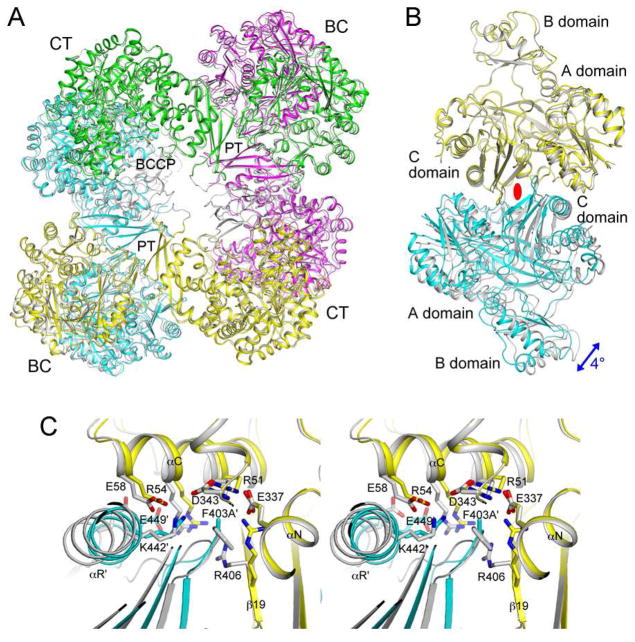
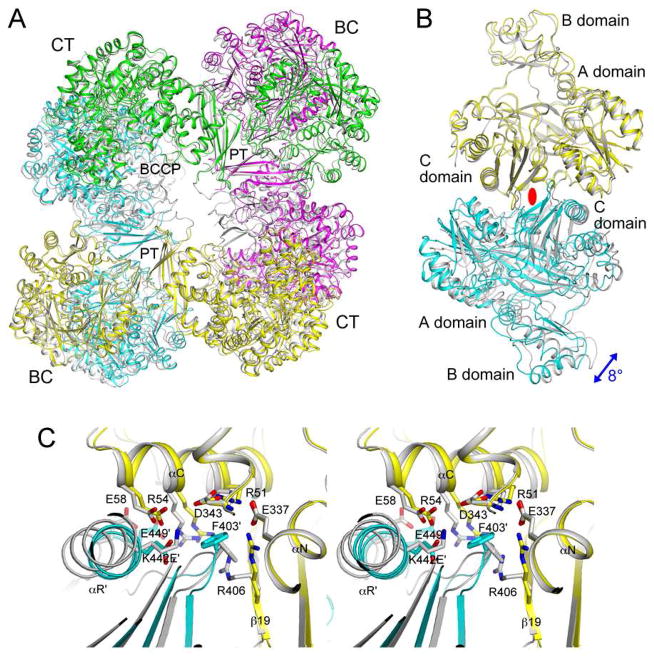
Biotin carboxylase (BC) is a conserved component among biotin-dependent carboxylases and catalyzes the MgATP-dependent carboxylation of biotin, using bicarbonate as the CO₂ donor. Studies with Escherichia coli BC have suggested long-range communication between the two active sites of a dimer, although its mechanism is not well understood. In addition, mutations in the dimer interface can produce stable monomers that are still catalytically active. A homologous dimer for the BC domain is observed in the structure of the tetrameric pyruvate carboxylase (PC) holoenzyme. We have introduced site-specific mutations into the BC domain dimer interface of Staphylococcus aureus PC (SaPC), equivalent to those used for E. coli BC, and also made chimeras replacing the SaPC BC domain with the E. coli BC subunit (EcBC chimera) or the yeast ACC BC domain (ScBC chimera). We assessed the catalytic activities of these mutants and characterized their oligomerization states by gel filtration and analytical ultracentrifugation experiments. The K442E mutant and the ScBC chimera disrupted the BC dimer and were catalytically inactive, while the F403A mutant and the EcBC chimera were still tetrameric and retained catalytic activity. The R54E mutant was also tetrameric but was catalytically inactive. Crystal structures of the R54E, F403A, and K442E mutants showed that they were tetrameric in the crystal, with conformational changes near the mutation site as well as in the tetramer organization. We have also produced the isolated BC domain of SaPC. In contrast to E. coli BC, the SaPC BC domain is monomeric in solution and catalytically inactive.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
