

Structural and functional characteristics of cGMP-dependent methionine oxidation in Arabidopsis thaliana proteins
- PMID: 23289948
- PMCID: PMC3544604
- DOI: 10.1186/1478-811X-11-1
Structural and functional characteristics of cGMP-dependent methionine oxidation in Arabidopsis thaliana proteins
Abstract
Background: Increasing structural and biochemical evidence suggests that post-translational methionine oxidation of proteins is not just a result of cellular damage but may provide the cell with information on the cellular oxidative status. In addition, oxidation of methionine residues in key regulatory proteins, such as calmodulin, does influence cellular homeostasis. Previous findings also indicate that oxidation of methionine residues in signaling molecules may have a role in stress responses since these specific structural modifications can in turn change biological activities of proteins.
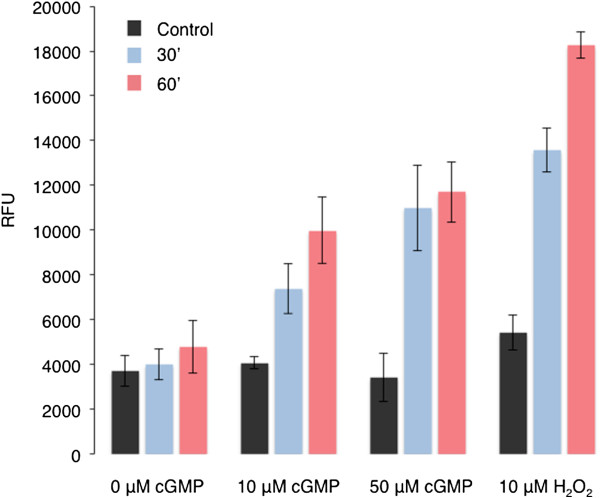
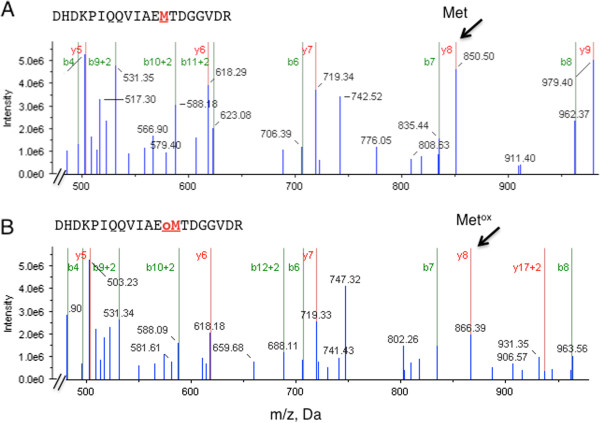
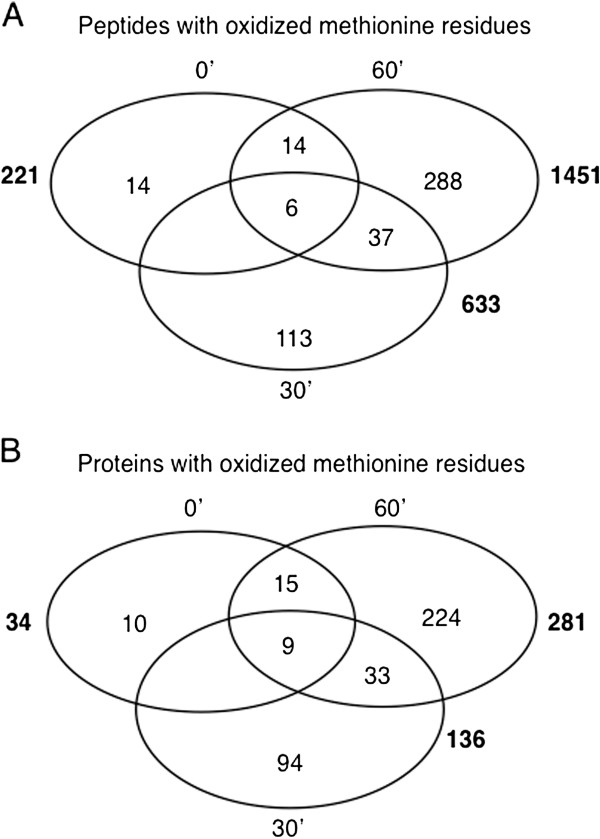
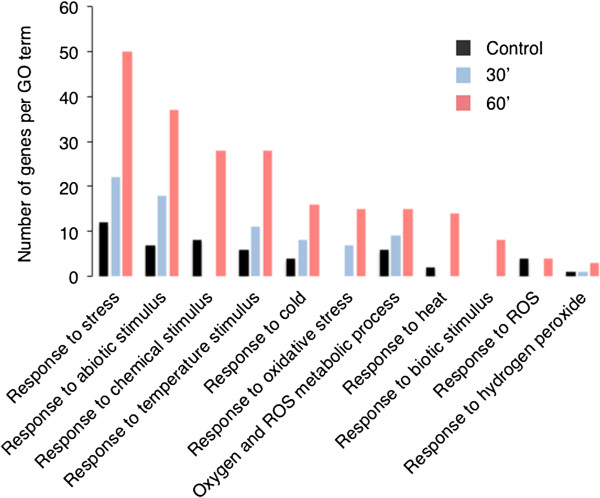
Findings: Here we use tandem mass spectrometry-based proteomics to show that treatment of Arabidopsis thaliana cells with a non-oxidative signaling molecule, the cell-permeant second messenger analogue, 8-bromo-3,5-cyclic guanosine monophosphate (8-Br-cGMP), results in a time-dependent increase in the content of oxidised methionine residues. Interestingly, the group of proteins affected by cGMP-dependent methionine oxidation is functionally enriched for stress response proteins. Furthermore, we also noted distinct signatures in the frequency of amino acids flanking oxidised and un-oxidised methionine residues on both the C- and N-terminus.
Conclusions: Given both a structural and functional bias in methionine oxidation events in response to a signaling molecule, we propose that these are indicative of a specific role of such post-translational modifications in the direct or indirect regulation of cellular responses. The mechanisms that determine the specificity of the modifications remain to be elucidated.
Figures
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
