

Mechanism and regulation of Cdc25/Twine protein destruction in embryonic cell-cycle remodeling
- PMID: 23290551
- PMCID: PMC4091758
- DOI: 10.1016/j.cub.2012.11.036
Mechanism and regulation of Cdc25/Twine protein destruction in embryonic cell-cycle remodeling
Abstract
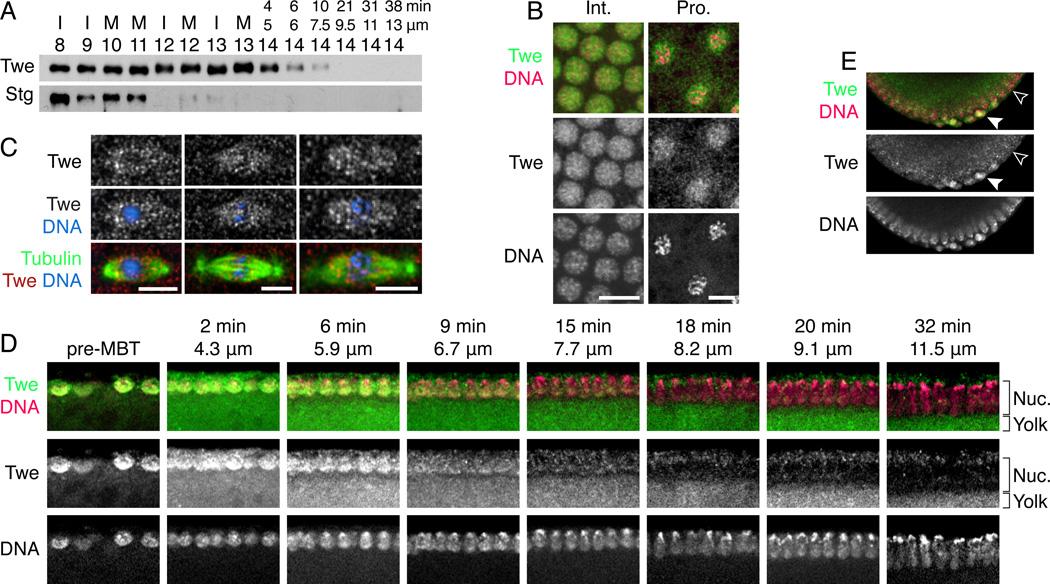
Background: In Drosophila embryos, the midblastula transition (MBT) dramatically remodels the cell cycle during the 14(th) interphase. Before the MBT, each cycle is composed of only a short S phase and mitosis. At the MBT, S phase is dramatically lengthened by the onset of late replication, and a G2 phase is introduced. Both changes set the stage for gastrulation and require downregulation of Cdc25 phosphatase, which was previously attributed to the elimination of its transcripts at the MBT.
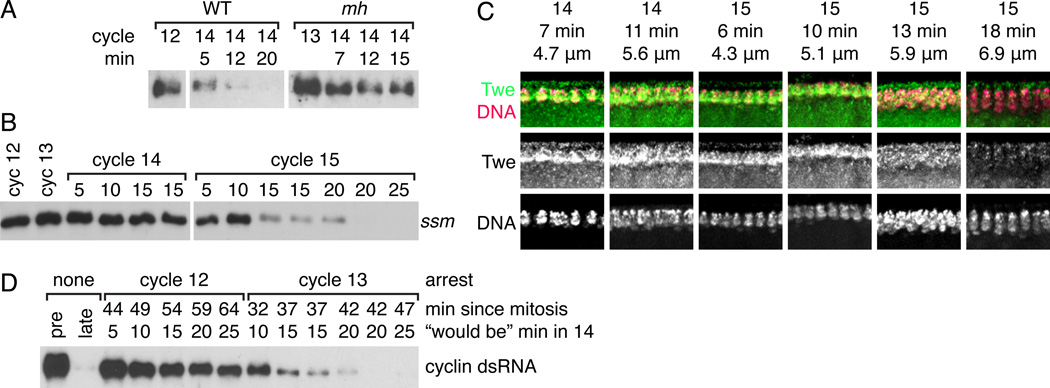
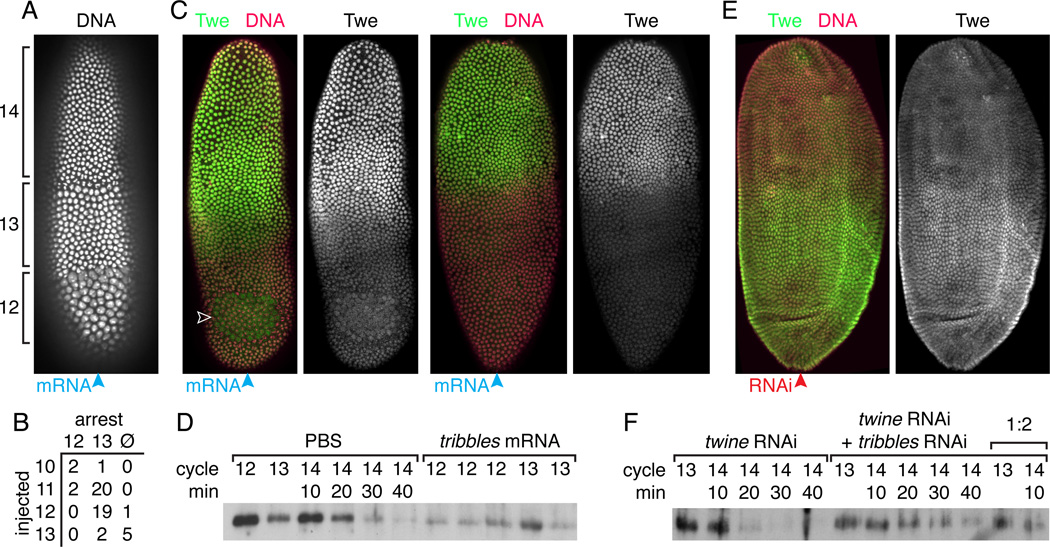
Results: Premature removal of cdc25 transcripts by RNAi did not affect progression to the MBT. Instead, an antibody against the Cdc25 isoform Twine showed that Twine protein was abundant and stable until the MBT, when it was destabilized and rapidly eliminated. Persistence of pre-MBT levels of Twine was sufficient to prevent cell-cycle slowing. Twine protein destruction was timed by the nucleocytoplasmic ratio and depended on the activation of zygotic transcription at the MBT, including expression of the gene tribbles, whose activity was sufficient to trigger Twine destruction and was required for prompt Twine disappearance.
Conclusions: We propose that the developmentally regulated destruction of Twine protein is a critical switch that contributes to the cell-cycle change at the MBT, including the addition of a G2 phase and onset of late replication. Moreover, we show that this destruction is triggered by the nucleocytoplasmic ratio-dependent onset of zygotic transcription of tribbles and other unknown genes.
Copyright © 2013 Elsevier Ltd. All rights reserved.
Figures



Comment in
-
Development: new wrinkles on genetic control of the MBT.Curr Biol. 2013 Jan 21;23(2):R65-7. doi: 10.1016/j.cub.2012.12.012. Curr Biol. 2013. PMID: 23347940
References
-
- Jeffrey PD, Russo AA, Polyak K, Gibbs E, Hurwitz J, Massagué J, Pavletich NP. Mechanism of CDK activation revealed by the structure of a cyclinA-CDK2 complex. Nature. 1995;376:313–320. - PubMed
-
- Edgar BA, Datar SA. Zygotic degradation of two maternal Cdc25 mRNAs terminates Drosophila's early cell cycle program. Genes Dev. 1996;10:1966–1977. - PubMed
-
- White-Cooper H, Alphey L, Glover DM. The cdc25 homologue twine is required for only some aspects of the entry into meiosis in Drosophila. J Cell Sci. 1993;106(Pt 4):1035–1044. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
