

Cotranslational response to proteotoxic stress by elongation pausing of ribosomes
- PMID: 23290916
- PMCID: PMC3570626
- DOI: 10.1016/j.molcel.2012.12.001
Cotranslational response to proteotoxic stress by elongation pausing of ribosomes
Abstract
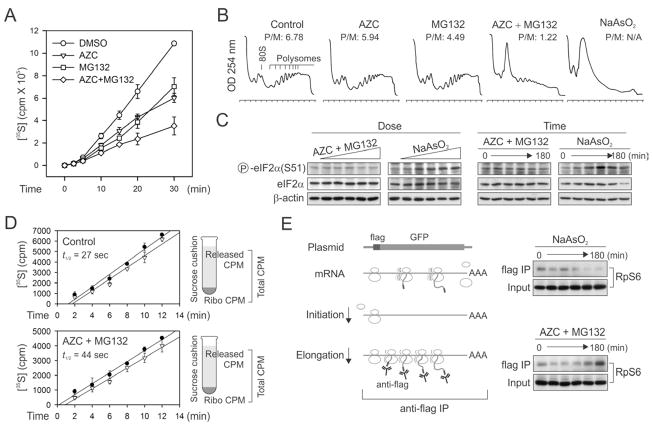
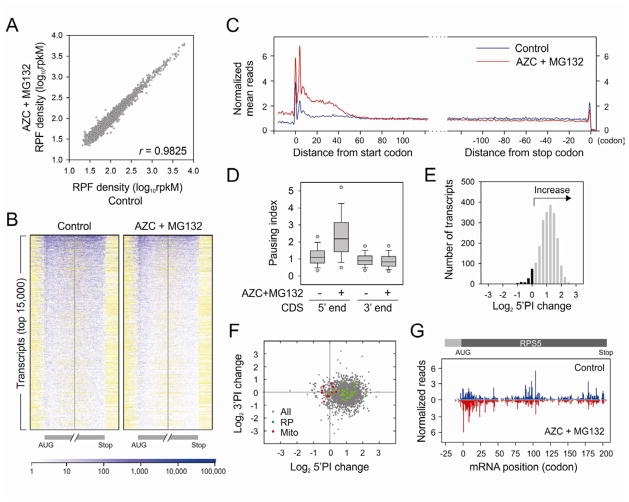
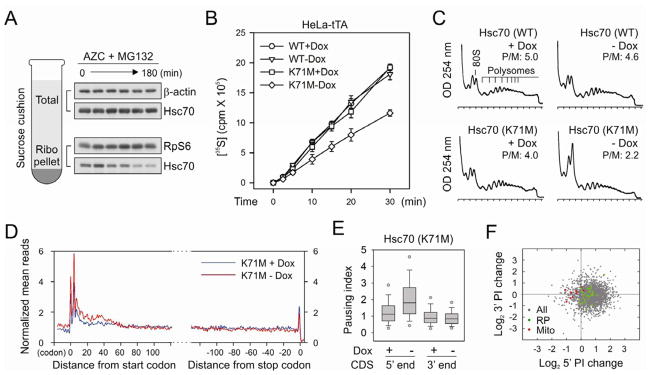
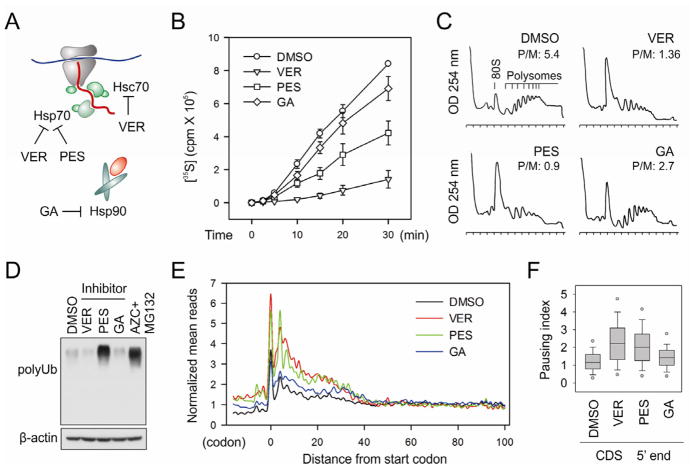
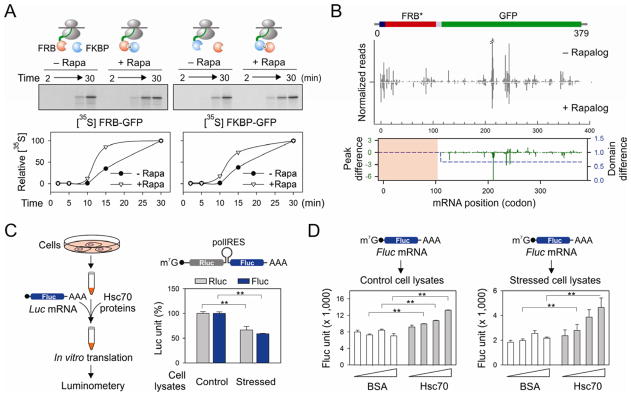
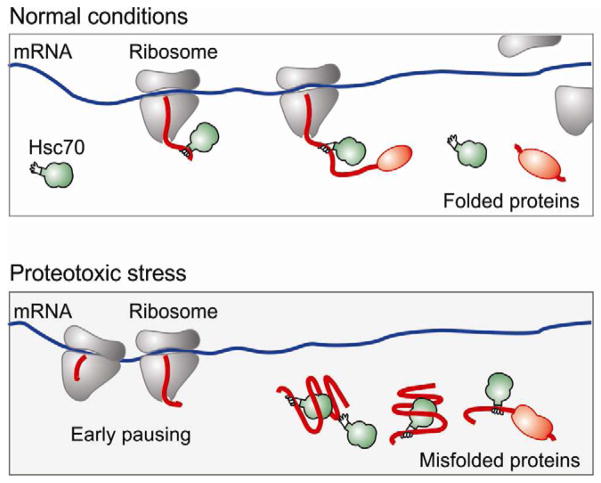
Translational control permits cells to respond swiftly to a changing environment. Rapid attenuation of global protein synthesis under stress conditions has been largely ascribed to the inhibition of translation initiation. Here we report that intracellular proteotoxic stress reduces global protein synthesis by halting ribosomes on transcripts during elongation. Deep sequencing of ribosome-protected messenger RNA (mRNA) fragments reveals an early elongation pausing, roughly at the site where nascent polypeptide chains emerge from the ribosomal exit tunnel. Inhibiting endogenous chaperone molecules by a dominant-negative mutant or chemical inhibitors recapitulates the early elongation pausing, suggesting a dual role of molecular chaperones in facilitating polypeptide elongation and cotranslational folding. Our results further support the chaperone "trapping" mechanism in promoting the passage of nascent chains. Our study reveals that translating ribosomes fine tune the elongation rate by sensing the intracellular folding environment. The early elongation pausing represents a cotranslational stress response to maintain the intracellular protein homeostasis.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
References
-
- Albanese V, Yam AY, Baughman J, Parnot C, Frydman J. Systems analyses reveal two chaperone networks with distinct functions in eukaryotic cells. Cell. 2006;124:75–88. - PubMed
-
- Bukau B, Weissman J, Horwich A. Molecular chaperones and protein quality control. Cell. 2006;125:443–451. - PubMed
-
- Choi J, Chen J, Schreiber SL, Clardy J. Structure of the FKBP12-rapamycin complex interacting with the binding domain of human FRAP. Science. 1996;273:239–242. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
