

Combined action of nucleic acid-sensing Toll-like receptors and TLR11/TLR12 heterodimers imparts resistance to Toxoplasma gondii in mice
- PMID: 23290966
- PMCID: PMC3552114
- DOI: 10.1016/j.chom.2012.12.003
Combined action of nucleic acid-sensing Toll-like receptors and TLR11/TLR12 heterodimers imparts resistance to Toxoplasma gondii in mice
Abstract
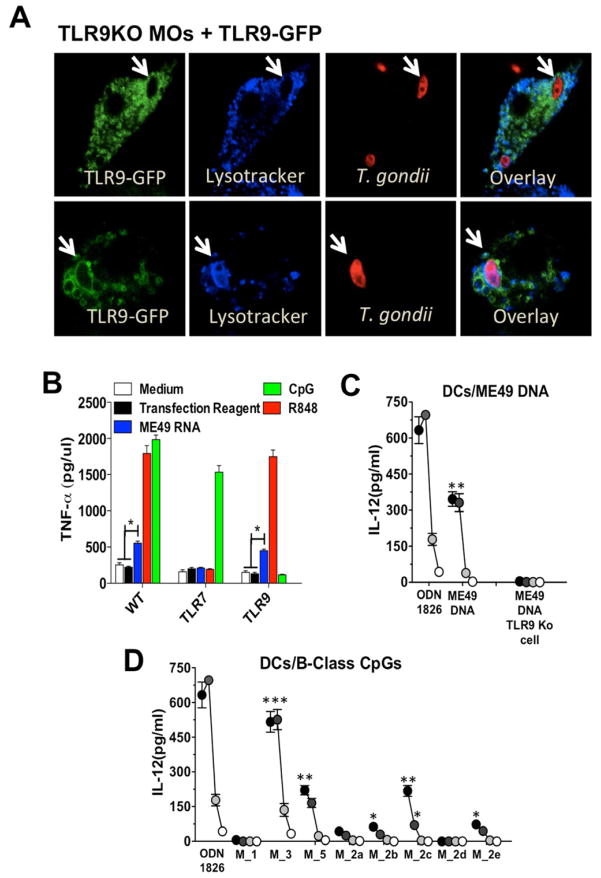
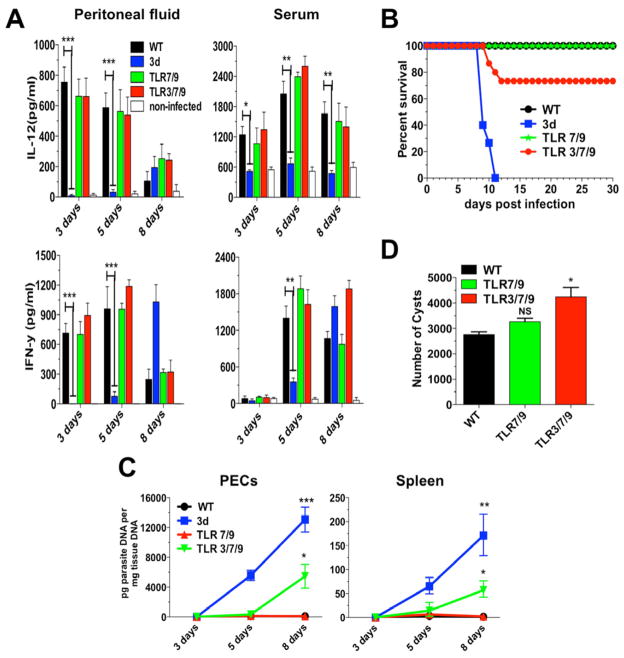
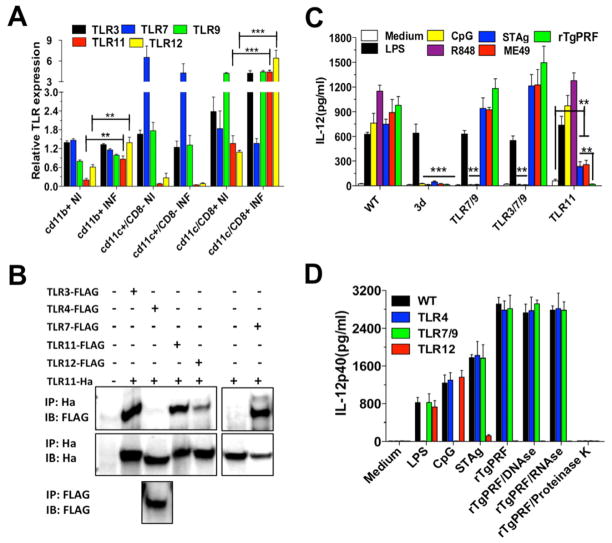
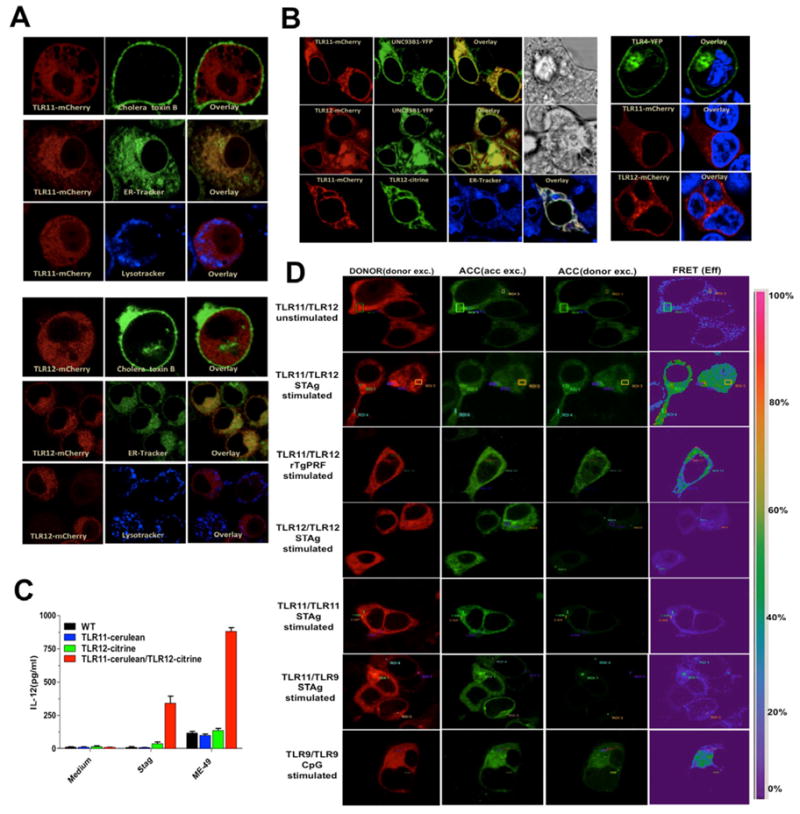
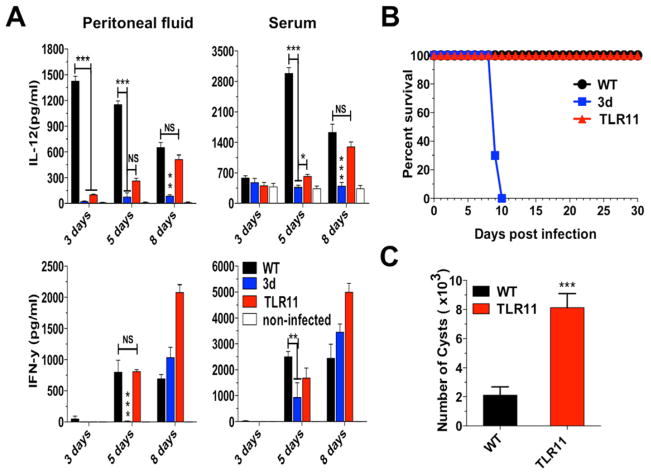
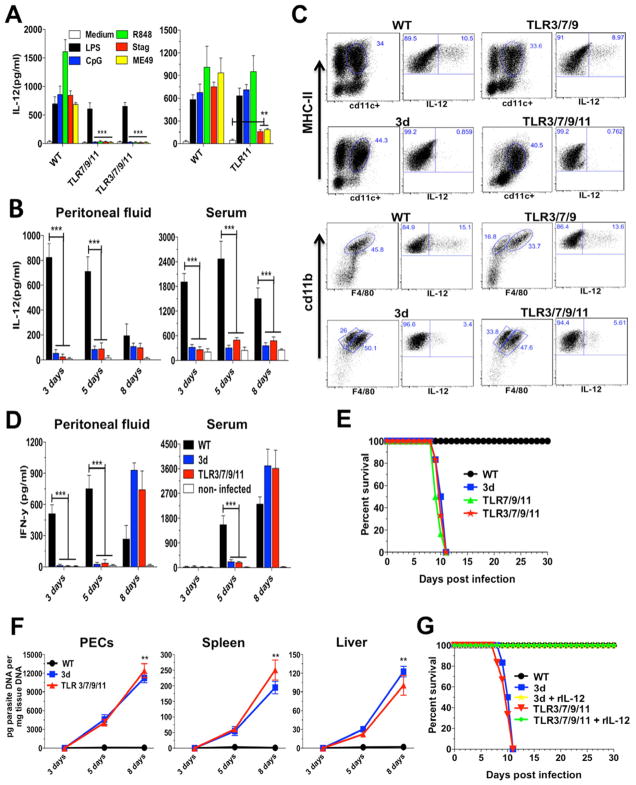
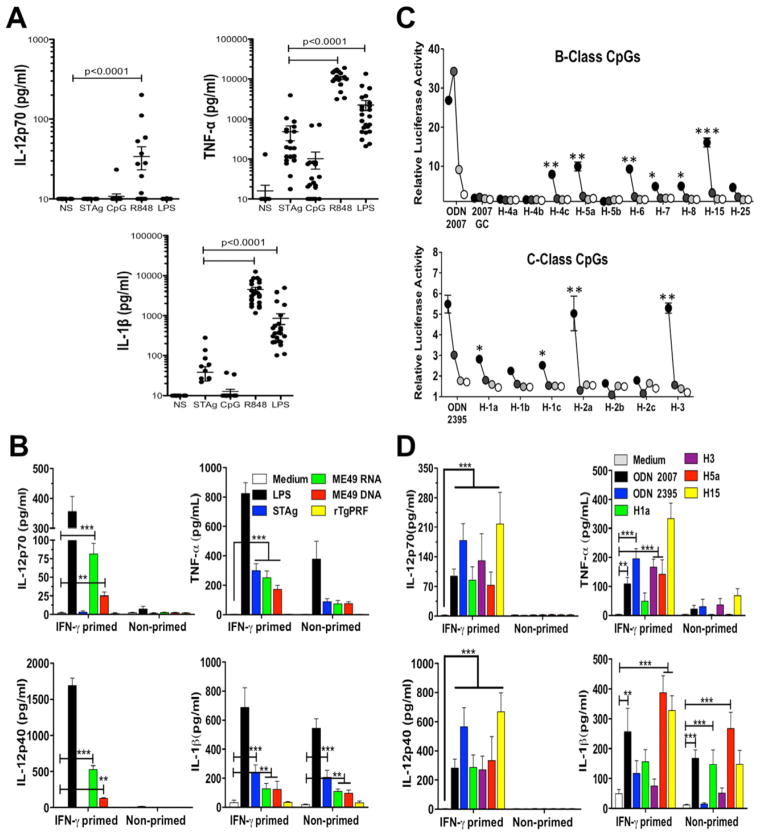
"Triple-defective" (3d) mice carrying a mutation in UNC93B1, a chaperone for the endosomal nucleic acid-sensing (NAS) Toll-like receptors TLR3, TLR7, and TLR9, are highly susceptible to Toxoplasma gondii infection. However, none of the single or even the triple NAS-TLR-deficient animals recapitulated the 3d susceptible phenotype to experimental toxoplasmosis. Investigating this further, we found that while parasite RNA and DNA activate innate immune responses via TLR7 and TLR9, TLR11 and TLR12 working as heterodimers are required for sensing and responding to Toxoplasma profilin. Consequently, the triple TLR7/TLR9/TLR11-deficient mice are highly susceptible to T. gondii infection, recapitulating the phenotype of 3d mice. Humans lack functional TLR11 and TLR12 genes. Consistently, human cells produce high levels of proinflammatory cytokines in response to parasite-derived RNA and DNA, but not to Toxoplasma profilin, supporting a more critical role for NAS-TLRs in human toxoplasmosis.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
References
-
- Alexopoulou L, Holt AC, Medzhitov R, Flavell RA. Recognition of double-stranded RNA and activation of NF-kappaB by Toll-like receptor 3. Nature. 2001;413:732–738. - PubMed
-
- Bartholomeu DC, Ropert C, Melo MB, Parroche P, Junqueira CF, Teixeira SM, Sirois C, Kasperkovitz P, Knetter CF, Lien E, et al. Recruitment and endo-lysosomal activation of TLR9 in dendritic cells infected with Trypanosoma cruzi. J Immunol. 2008;181:1333–1344. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
