

Dysfunctional potassium channel subunit interaction as a novel mechanism of long QT syndrome
- PMID: 23291057
- PMCID: PMC4770260
- DOI: 10.1016/j.hrthm.2012.12.033
Dysfunctional potassium channel subunit interaction as a novel mechanism of long QT syndrome
Abstract
Background: The slowly-activating delayed rectifier current IKs contributes to repolarization of the cardiac action potential, and is composed of a pore-forming α-subunit, KCNQ1, and a modulatory β-subunit, KCNE1. Mutations in either subunit can cause long QT syndrome, a potentially fatal arrhythmic disorder. How KCNE1 exerts its extensive control over the kinetics of IKs remains unresolved
Objective: To evaluate the impact of a novel KCNQ1 mutation on IKs channel gating and kinetics
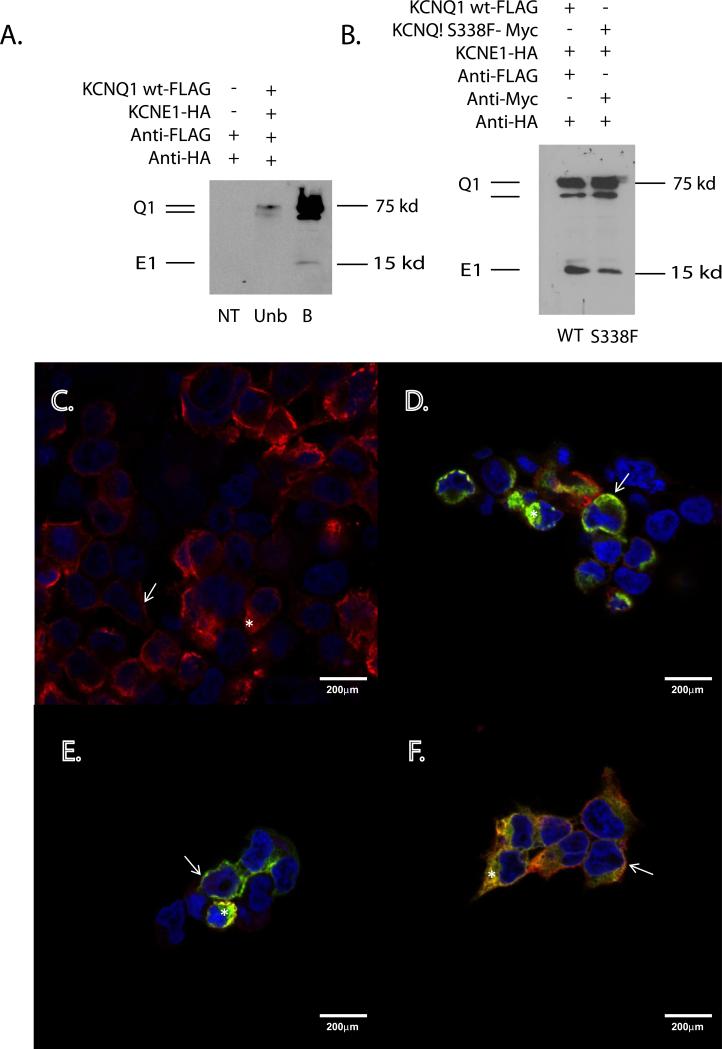
Methods: KCNQ1 mutations were expressed in Xenopus oocytes in the presence and absence of KCNE1. Voltage clamping and MODELLER software were used to characterize and model channel function. Mutant and wt genes were cloned into FLAG, Myc and HA expression vectors to achieve differential epitope tagging, and expressed in HEK293 cells for immunohistochemical localization and surface biotinylation assay.
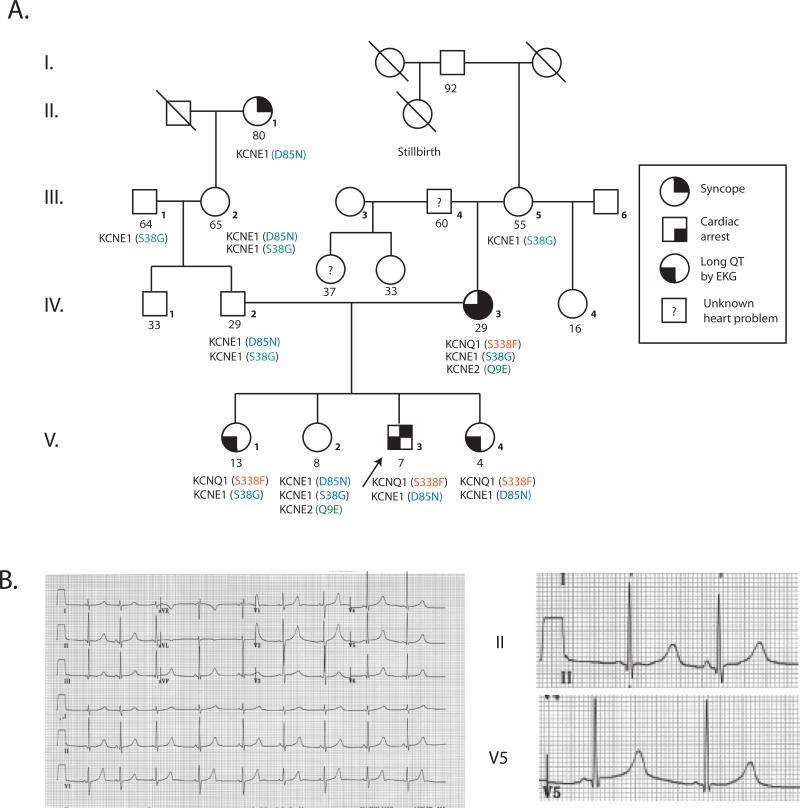
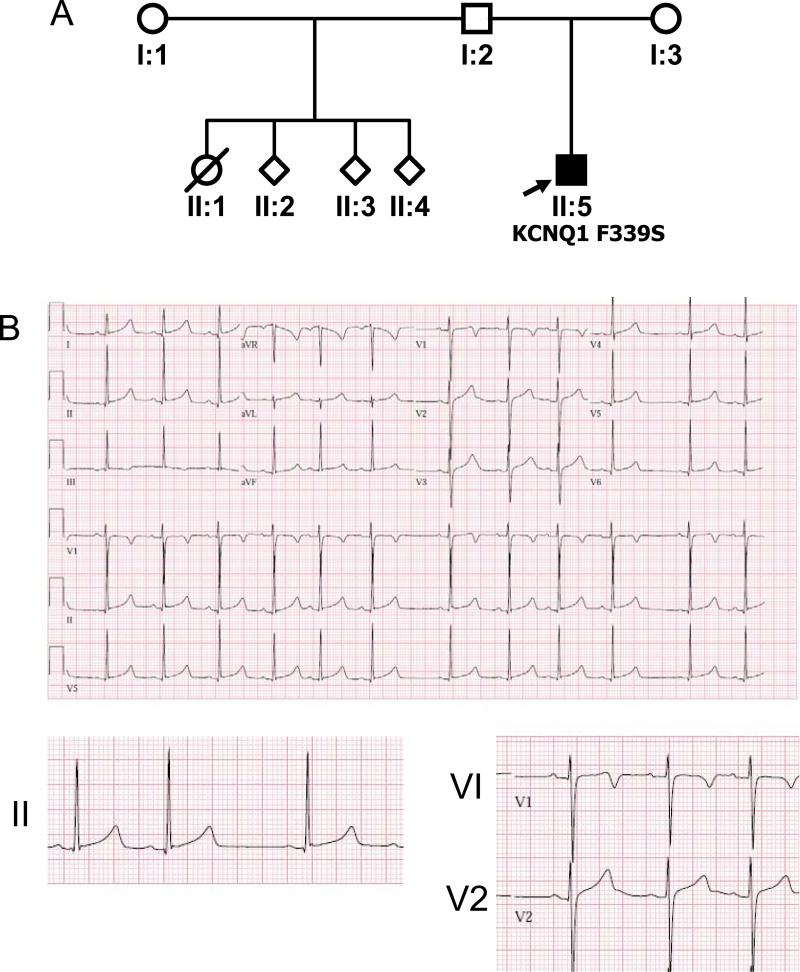
Results: We identified 2 adjacent mutations, S338F and F339S, in the KCNQ1 S6 domain in unrelated probands. The novel KCNQ1 S338F mutation segregated with prolonged QT interval and torsade de pointes; the second variant, F339S, was associated with fetal bradycardia and prolonged QT interval, but no other clinical events. S338F channels expressed in Xenopus oocytes had slightly increased peak conductance relative to wild type, with a more positive activation voltage. F339S channels conducted minimal current. Unexpectedly, S338F currents were abolished by co-expression with intact WT KCNE1 or its C-terminus (aa63-129), despite normal membrane trafficking and surface co-localization of KCNQ1 S338F and wt KCNE1. Structural modeling indicated that the S338F mutation specifically alters the interaction between the S6 domain of one KCNQ1 subunit and the S4-S5 linker of another, inhibiting voltage-induced movement synergistically with KCNE1 binding.
Conclusions: A novel KCNQ1 mutation specifically impaired channel function in the presence of KCNE1. Our structural model shows that this mutation effectively immobilizes voltage gating by an inhibitory interaction that is additive with that of KCNE1. Our findings illuminate a previously unreported mechanism for LQTS, and validate recent theoretical models of the closed state of the KCNQ1:KCNE1 complex.
Copyright © 2013 Heart Rhythm Society. Published by Elsevier Inc. All rights reserved.
Figures




Comment in
-
Ion channel complex disease in long QT syndrome.Heart Rhythm. 2013 May;10(5):738-9. doi: 10.1016/j.hrthm.2013.01.026. Epub 2013 Jan 23. Heart Rhythm. 2013. PMID: 23354076 No abstract available.
References
-
- Modell SM, Lehmann MH. The long QT syndrome family of cardiac ion channelopathies: a HuGE review. Genet Med. 2006;8:143–155. - PubMed
-
- Sanguinetti MC, Curran ME, Zou A, et al. Coassembly of K(V)LQT1 and minK (IsK) proteins to form cardiac I(Ks) potassium channel. Nature. 1996;384:80–83. - PubMed
-
- Barhanin J, Lesage F, Guillemare E, Fink M, Lazdunski M, Romey G. K(V)LQT1 and lsK (minK) proteins associate to form the I(Ks) cardiac potassium current. Nature. 1996;384:78–80. - PubMed
-
- Nerbonne JM, Kass RS. Molecular physiology of cardiac repolarization. Physiol Rev. 2005;85:1205–1253. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
