

RPA accumulation during class switch recombination represents 5'-3' DNA-end resection during the S-G2/M phase of the cell cycle
- PMID: 23291097
- PMCID: PMC3563767
- DOI: 10.1016/j.celrep.2012.12.006
RPA accumulation during class switch recombination represents 5'-3' DNA-end resection during the S-G2/M phase of the cell cycle
Abstract
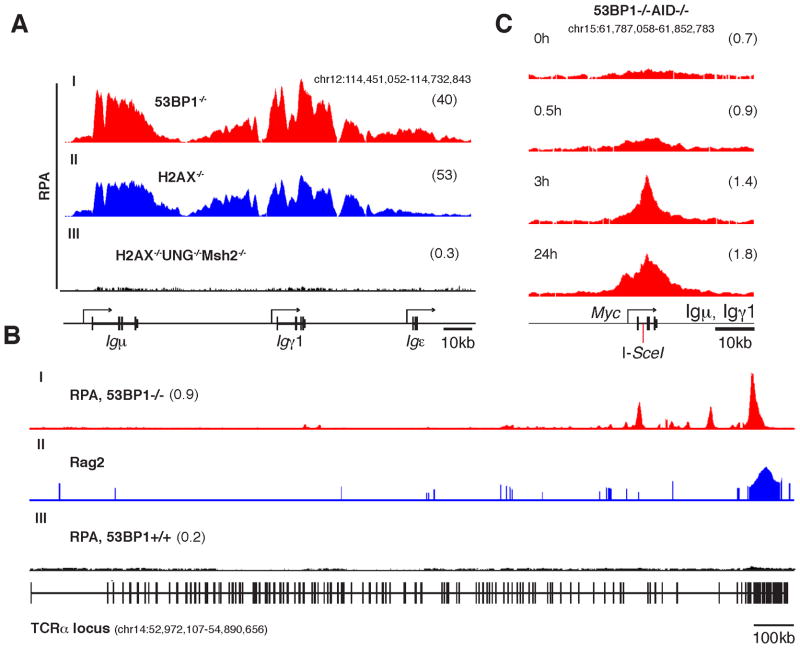
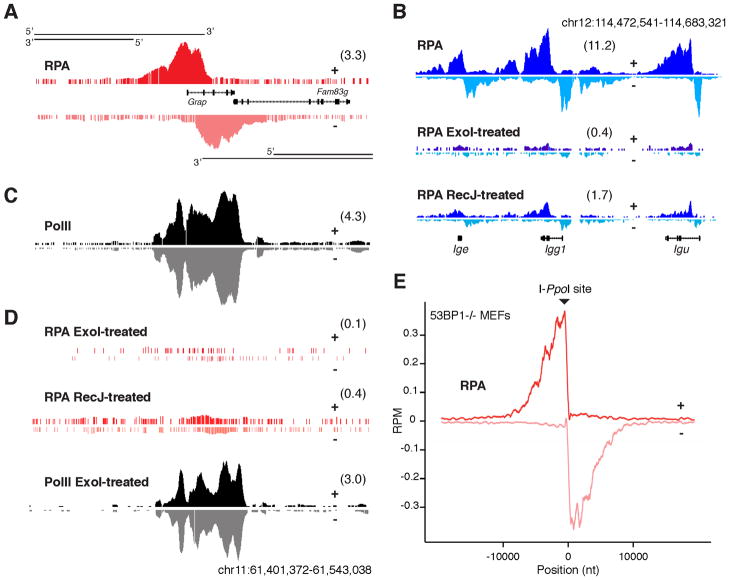
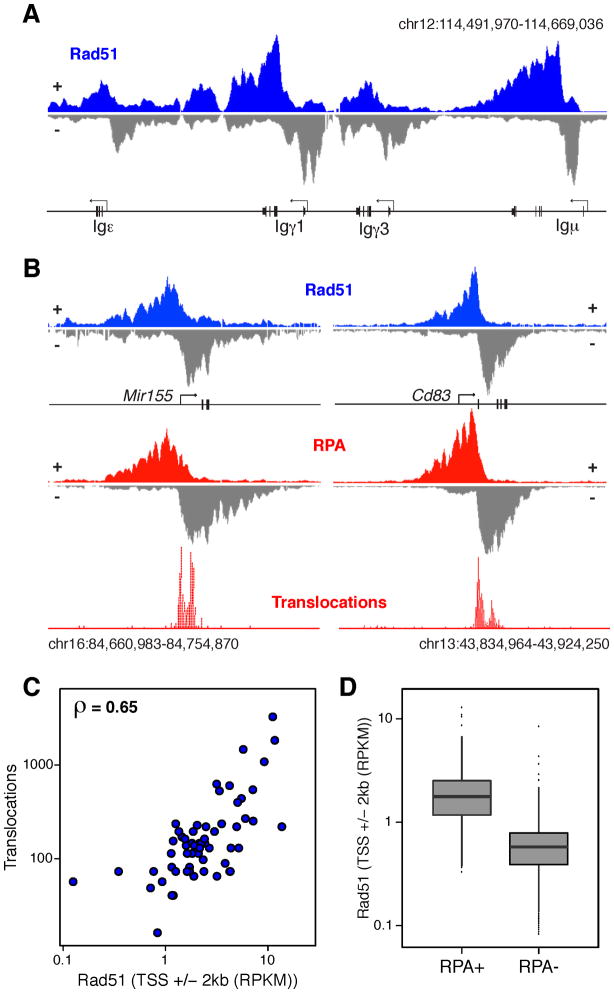
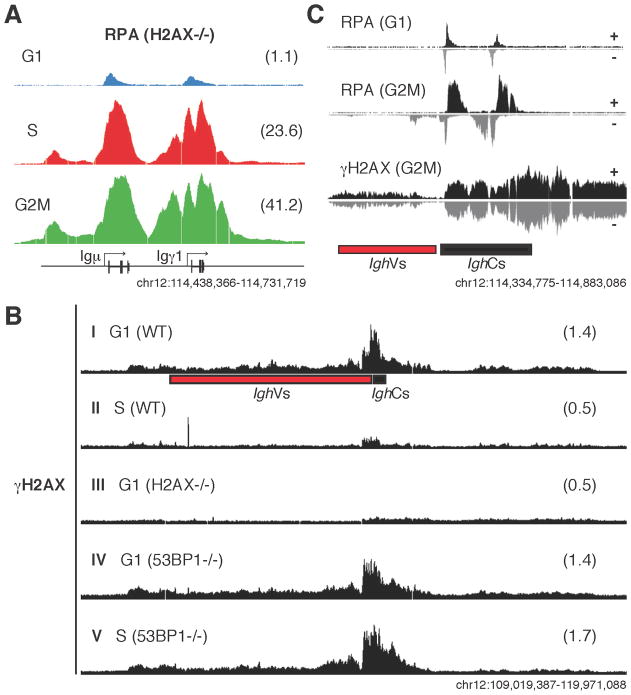
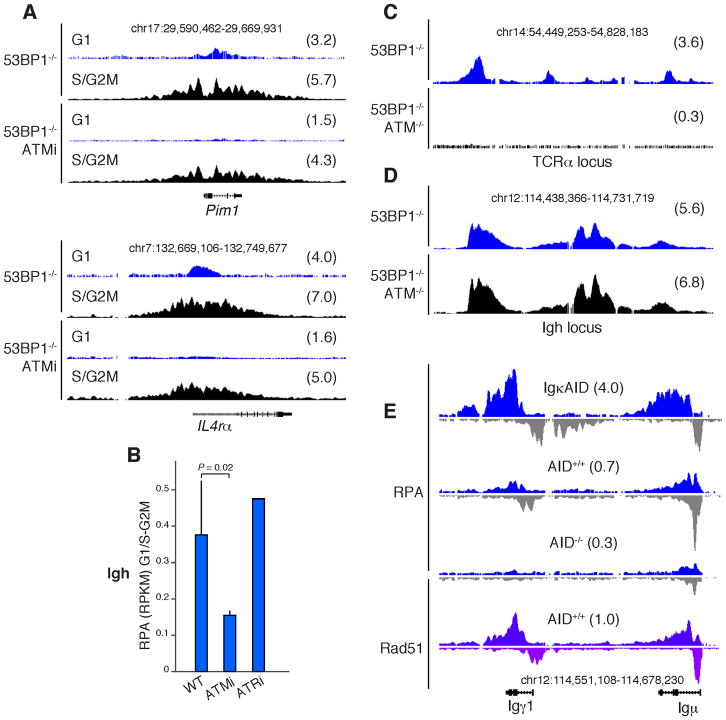
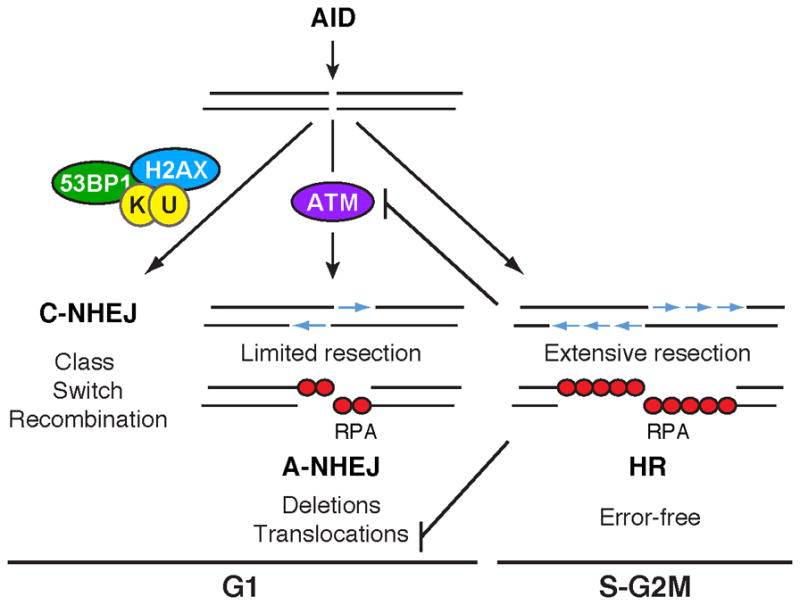
Activation-induced cytidine deaminase (AID) promotes chromosomal translocations by inducing DNA double-strand breaks (DSBs) at immunoglobulin (Ig) genes and oncogenes in the G1 phase. RPA is a single-stranded DNA (ssDNA)-binding protein that associates with resected DSBs in the S phase and facilitates the assembly of factors involved in homologous repair (HR), such as Rad51. Notably, RPA deposition also marks sites of AID-mediated damage, but its role in Ig gene recombination remains unclear. Here, we demonstrate that RPA associates asymmetrically with resected ssDNA in response to lesions created by AID, recombination-activating genes (RAG), or other nucleases. Small amounts of RPA are deposited at AID targets in G1 in an ATM-dependent manner. In contrast, recruitment in the S-G2/M phase is extensive, ATM independent, and associated with Rad51 accumulation. In the S-G2/M phase, RPA increases in nonhomologous-end-joining-deficient lymphocytes, where there is more extensive DNA-end resection. Thus, most RPA recruitment during class switch recombination represents salvage of unrepaired breaks by homology-based pathways during the S-G2/M phase of the cell cycle.
Copyright © 2013 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Bakkenist CJ, Kastan MB. DNA damage activates ATM through intermolecular autophosphorylation and dimer dissociation. Nature. 2003;421:499–506. - PubMed
-
- Barlow C, Hirotsune S, Paylor R, Liyanage M, Eckhaus M, Collins F, Shiloh Y, Crawley JN, Ried T, Tagle D, et al. Atm-deficient mice: a paradigm of ataxia telangiectasia. Cell. 1996;86:159–171. - PubMed
-
- Bassing CH, Suh H, Ferguson DO, Chua KF, Manis J, Eckersdorff M, Gleason M, Bronson R, Lee C, Alt FW. Histone H2AX: a dosage-dependent suppressor of oncogenic translocations and tumors. Cell. 2003;114:359–370. - PubMed
-
- Bishop DK, Park D, Xu L, Kleckner N. DMC1: a meiosis-specific yeast homolog of E. coli recA required for recombination, synaptonemal complex formation, and cell cycle progression. Cell. 1992;69:439–456. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
