

Under lock and key: spatiotemporal regulation of WASP family proteins coordinates separate dynamic cellular processes
- PMID: 23291261
- PMCID: PMC3656410
- DOI: 10.1016/j.semcdb.2012.12.005
Under lock and key: spatiotemporal regulation of WASP family proteins coordinates separate dynamic cellular processes
Abstract
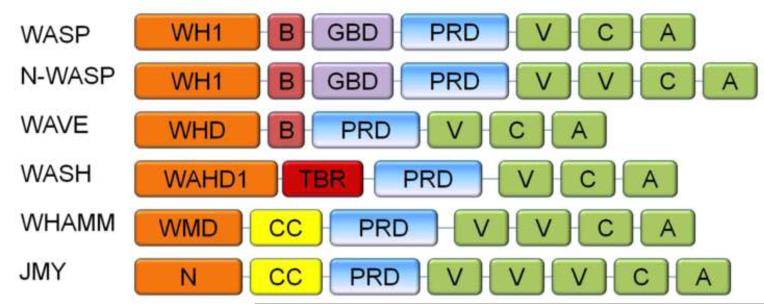
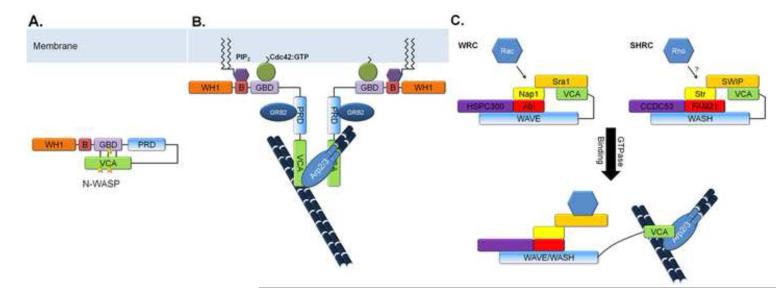
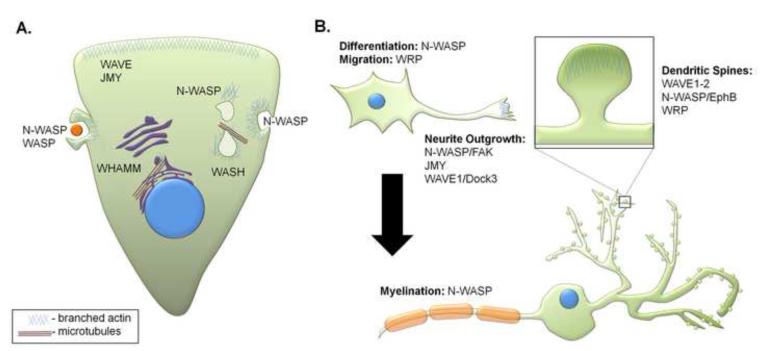
WASP family proteins are nucleation promoting factors that bind to and activate the Arp2/3 complex in order to stimulate nucleation of branched actin filaments. The WASP family consists of WASP, N-WASP, WAVE1-3, WASH, and the novel family members WHAMM and JMY. Each of the family members contains a C-terminus responsible for their nucleation promoting activity and unique N-termini that allow for them to be regulated in a spatiotemporal manner. Upon activation they reorganize the cytoskeleton for different cellular functions depending on their subcellular localization and regulatory protein interactions. Emerging evidence indicates that WASH, WHAMM, and JMY have functions that require the coordination of both actin polymerization and microtubule dynamics. Here, we review the mechanisms of regulation for each family member and their associated in vivo functions including cell migration, vesicle trafficking, and neuronal development.
Copyright © 2013 Elsevier Ltd. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
