

Recoding RNA editing of AZIN1 predisposes to hepatocellular carcinoma
- PMID: 23291631
- PMCID: PMC3783260
- DOI: 10.1038/nm.3043
Recoding RNA editing of AZIN1 predisposes to hepatocellular carcinoma
Abstract
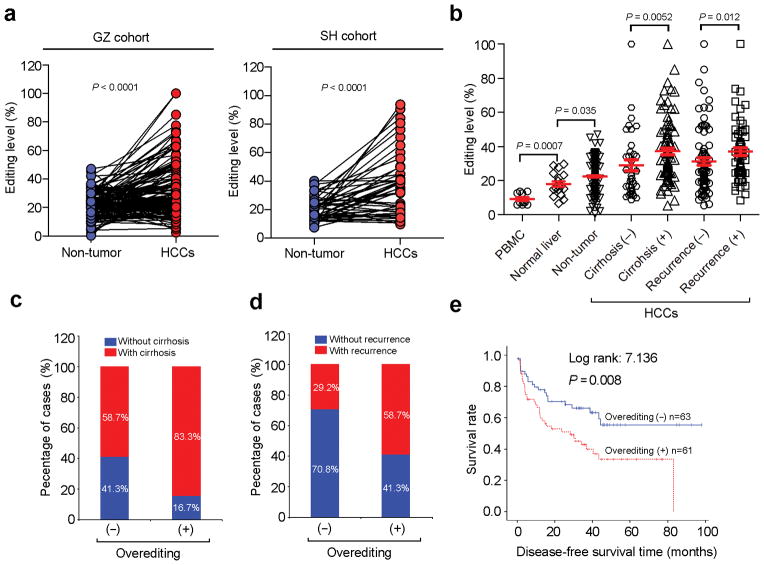
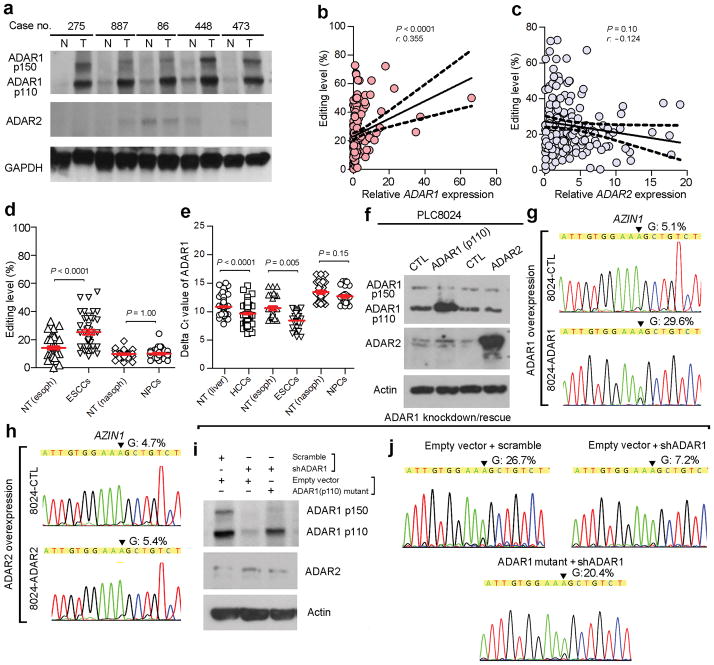
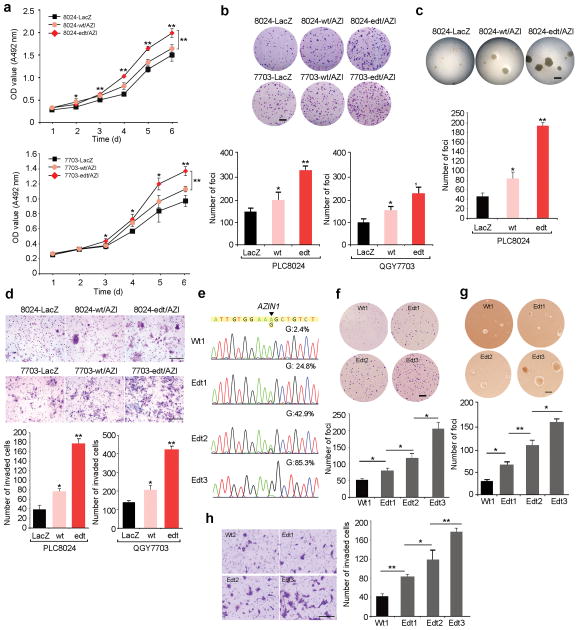
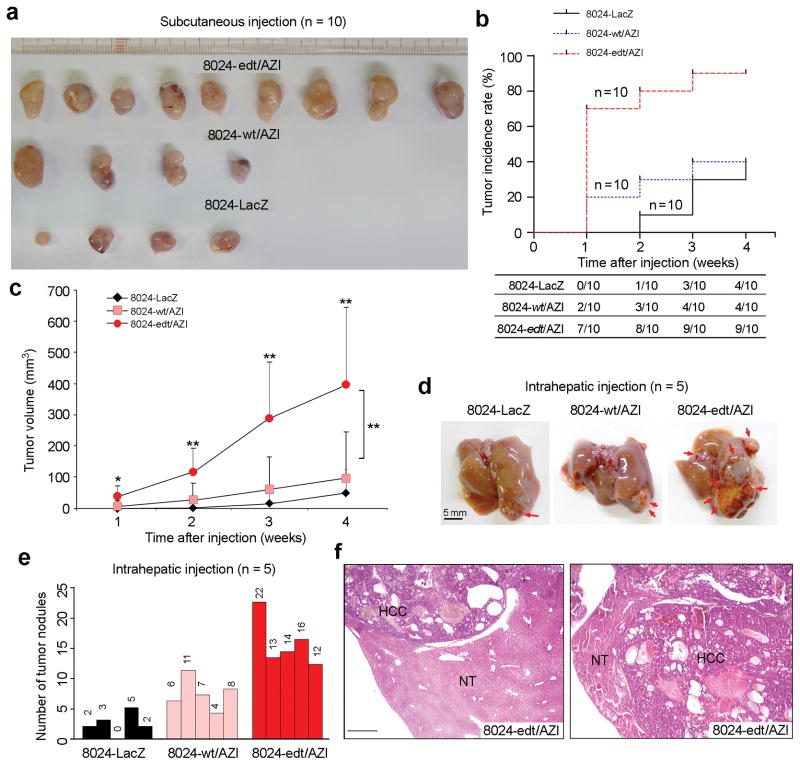
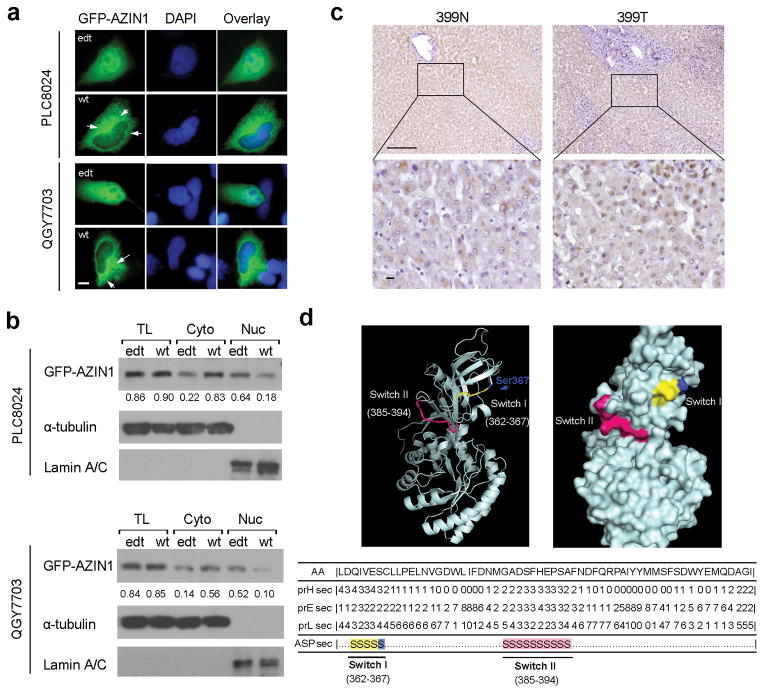
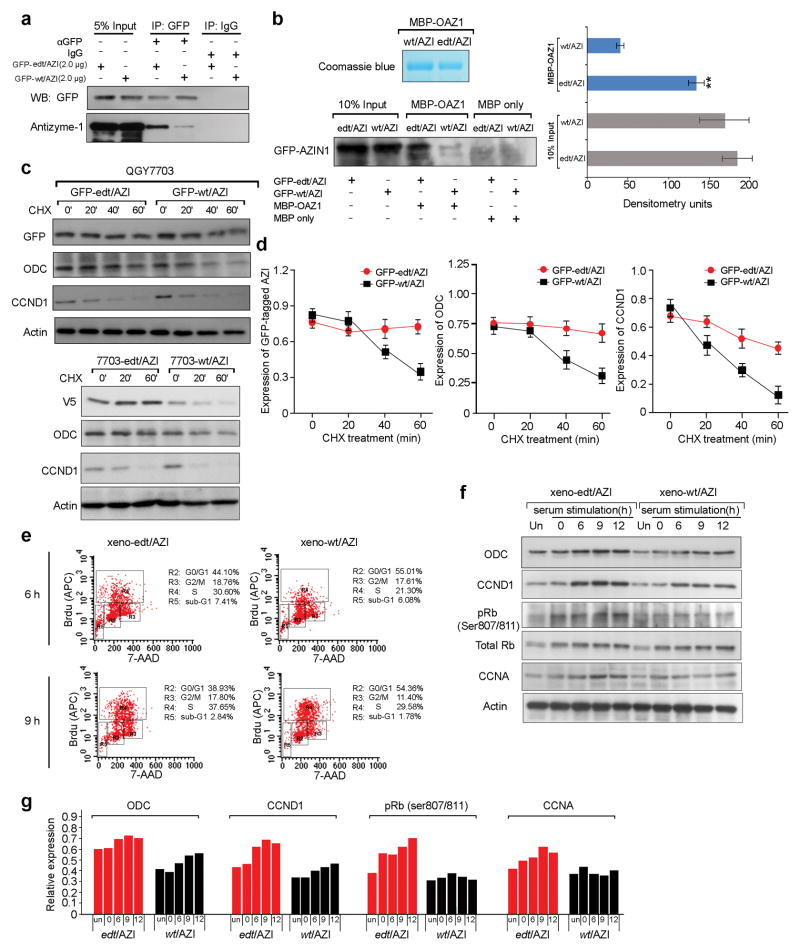
A better understanding of human hepatocellular carcinoma (HCC) pathogenesis at the molecular level will facilitate the discovery of tumor-initiating events. Transcriptome sequencing revealed that adenosine-to-inosine (A→I) RNA editing of AZIN1 (encoding antizyme inhibitor 1) is increased in HCC specimens. A→I editing of AZIN1 transcripts, specifically regulated by ADAR1 (encoding adenosine deaminase acting on RNA-1), results in a serine-to-glycine substitution at residue 367 of AZIN1, located in β-strand 15 (β15) and predicted to cause a conformational change, induced a cytoplasmic-to-nuclear translocation and conferred gain-of-function phenotypes that were manifested by augmented tumor-initiating potential and more aggressive behavior. Compared with wild-type AZIN1 protein, the edited form has a stronger affinity to antizyme, and the resultant higher AZIN1 protein stability promotes cell proliferation through the neutralization of antizyme-mediated degradation of ornithine decarboxylase (ODC) and cyclin D1 (CCND1). Collectively, A→I RNA editing of AZIN1 may be a potential driver in the pathogenesis of human cancers, particularly HCC.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Comment in
-
RNA editing enters the limelight in cancer.Nat Med. 2013 Feb;19(2):130-1. doi: 10.1038/nm.3072. Nat Med. 2013. PMID: 23389604 No abstract available.
-
Recoding RNA editing of AZIN1 is oncogenic in hepatocellular carcinoma.Cancer Discov. 2013 Mar;3(3):OF12. doi: 10.1158/2159-8290.CD-RW2013-017. Epub 2013 Jan 24. Cancer Discov. 2013. PMID: 23475882
References
-
- Parkin DM, Bray F, Ferlay J, Pisani P. Global cancer statistics, 2002. CA Cancer J Clin. 2005;55:74–108. - PubMed
-
- Farazi PA, DePinho RA. Hepatocellular carcinoma pathogenesis: from genes to environment. Nat Rev Cancer. 2006;6:674–687. - PubMed
-
- Thorgeirsson SS, Grisham JW. Molecular pathogenesis of human hepatocellular carcinoma. Nat Genet. 2002;31:339–346. - PubMed
-
- Meyerson M, Gabriel S, Getz G. Advances in understanding cancer genomes through second-generation sequencing. Nat Rev Genet. 11:685–696. - PubMed
-
- Morozova O, Marra MA. Applications of next-generation sequencing technologies in functional genomics. Genomics. 2008;92:255–264. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
