

Phenotype and function of B cells and dendritic cells from interferon regulatory factor 5-deficient mice with and without a mutation in DOCK2
- PMID: 23291967
- PMCID: PMC3631000
- DOI: 10.1093/intimm/dxs114
Phenotype and function of B cells and dendritic cells from interferon regulatory factor 5-deficient mice with and without a mutation in DOCK2
Abstract
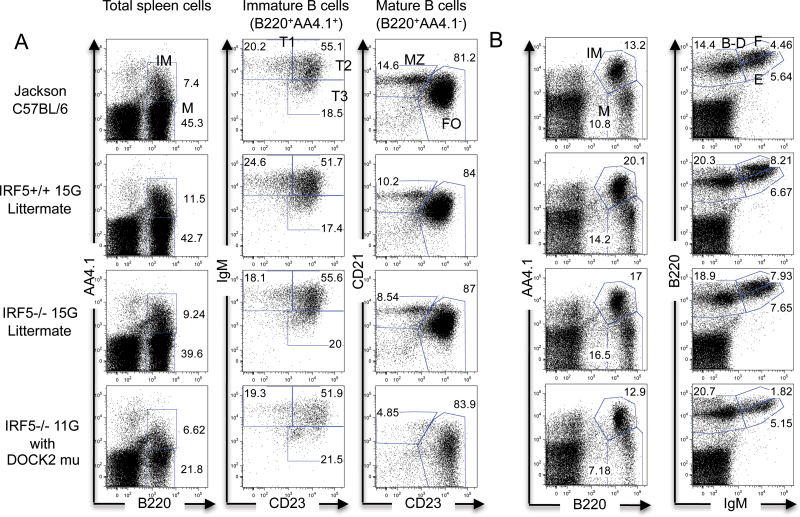
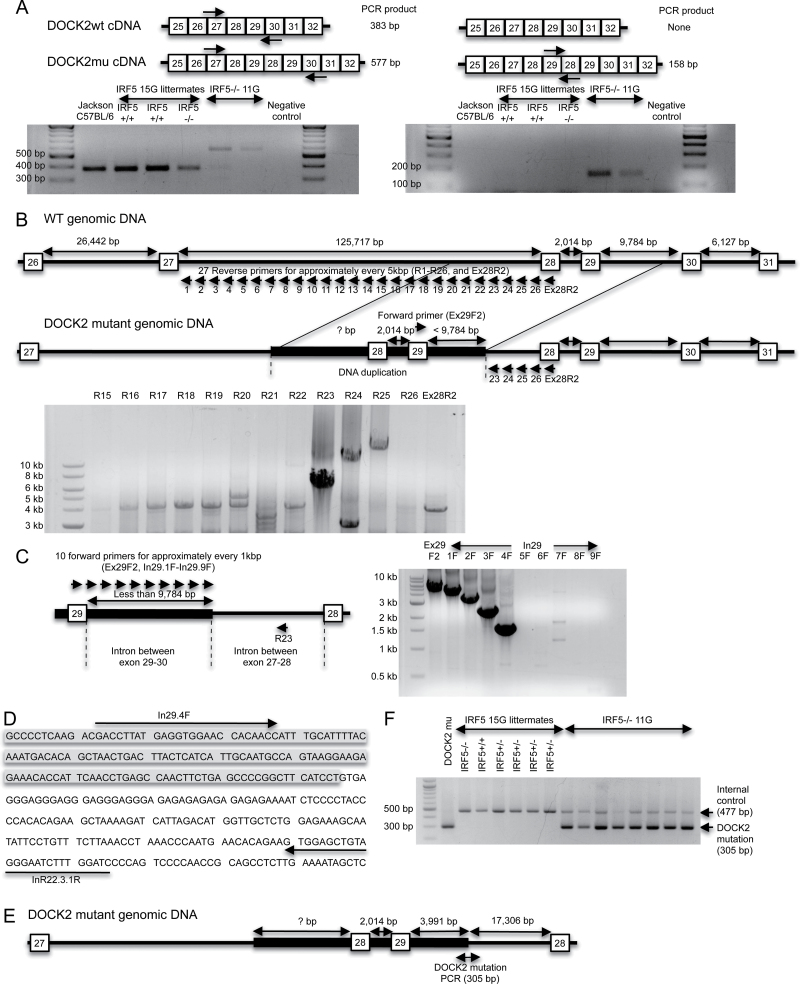
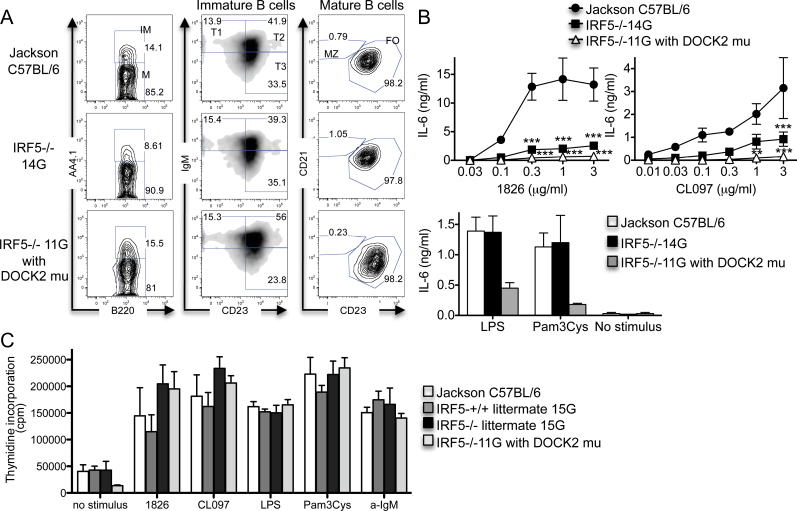
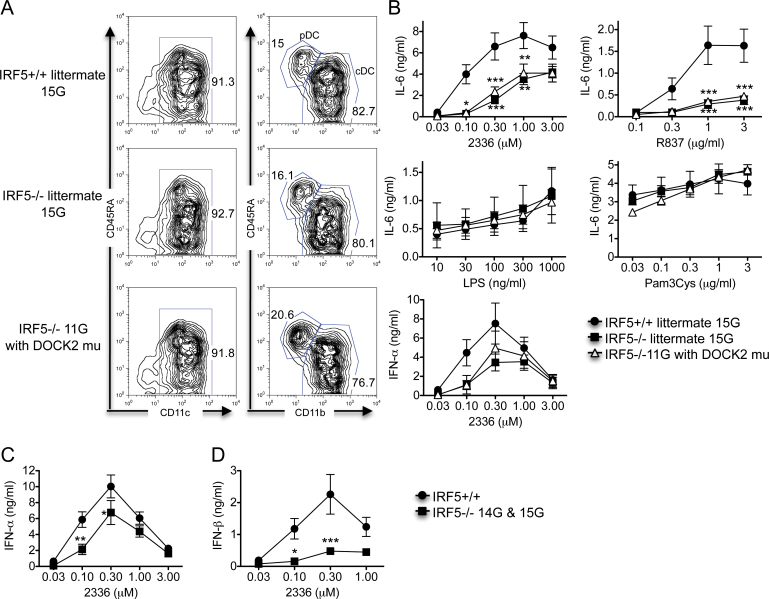
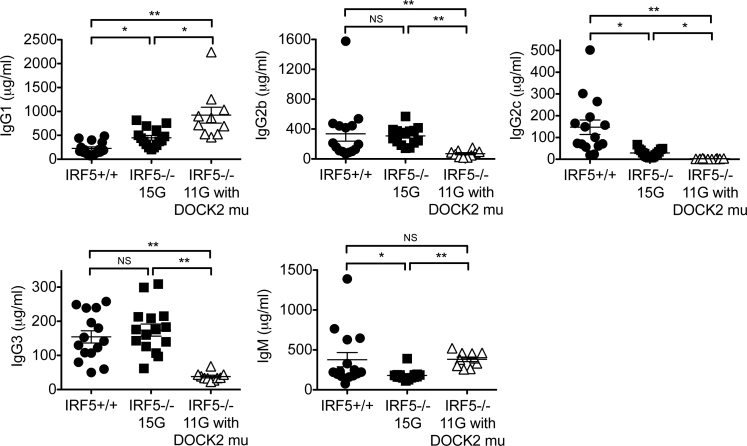
Interferon regulatory factor 5-deficient (IRF5 (-/-) ) mice have been used for many studies of IRF5 biology. A recent report identifies a mutation in dedicator of cytokinesis 2 (DOCK2) as being responsible for the abnormal B-cell development phenotype observed in the IRF5 (-/-) line. Both dedicator of cytokinesis 2 (DOCK2) and IRF5 play important roles in immune cell function, raising the issue of whether immune effects previously associated with IRF5 are due to IRF5 or DOCK2. Here, we defined the insertion end-point of the DOCK2 mutation and designed a novel PCR to detect the mutation in genomic DNA. We confirmed the association of the DOCK2 mutation and the abnormal B-cell phenotype in our IRF5 (-/-) line and also established another IRF5 (-/-) line without the DOCK2 mutation. These two lines were used to compare the role of IRF5 in dendritic cells (DCs) and B cells in the presence or absence of the DOCK2 mutation. IRF5 deficiency reduces IFN-α, IFN-β and IL-6 production by Toll-like receptor 9 (TLR9)- and TLR7-stimulated DCs and reduces TLR7- and TLR9-induced IL-6 production by B cells to a similar extent in the two lines. Importantly however, IRF5 (-/-) mice with the DOCK2 mutation have higher serum levels of IgG1 and lower levels of IgG2b, IgG2a/c and IgG3 than IRF5 (-/-) mice without the DOCK2 mutation, suggesting that the DOCK2 mutation confers additional Th2-type effects. Overall, these studies help clarify the function of IRF5 in B cells and DCs in the absence of the DOCK2 mutation. In addition, the PCR described will be useful for other investigators using the IRF5 (-/-) mouse line.
Figures
References
-
- Tamura T., Yanai H., Savitsky D., Taniguchi T. 2008. The IRF family transcription factors in immunity and oncogenesis. Annu. Rev. Immunol. 26: 535 - PubMed
-
- Barnes B. J., Moore P. A., Pitha P. M. 2001. Virus-specific activation of a novel interferon regulatory factor, IRF-5, results in the induction of distinct interferon alpha genes. J. Biol. Chem. 276: 23382 - PubMed
-
- Takaoka A., Yanai H., Kondo S., et al. 2005. Integral role of IRF-5 in the gene induction programme activated by Toll-like receptors. Nature 434: 243 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
