

Closing the circle of germline and stem cells: the Primordial Stem Cell hypothesis
- PMID: 23294912
- PMCID: PMC3599645
- DOI: 10.1186/2041-9139-4-2
Closing the circle of germline and stem cells: the Primordial Stem Cell hypothesis
Abstract
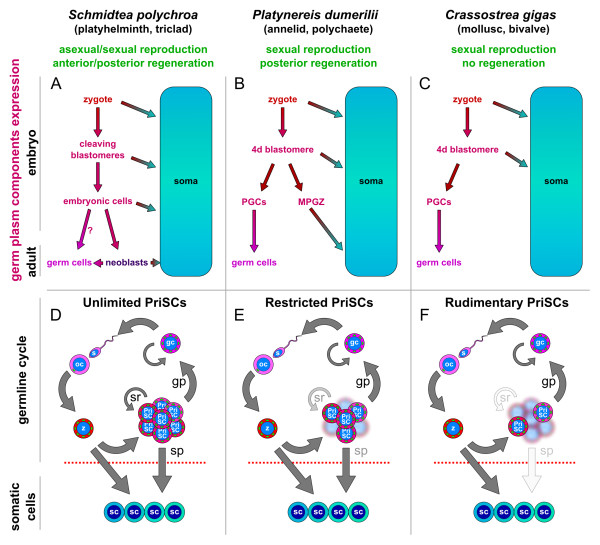
Background: Germline determination is believed to occur by either preformation or epigenesis. Animals that undergo germ cell specification by preformation have a continuous germline. However, animals with germline determination by epigenesis have a discontinuous germline, with somatic cells intercalated. This vision is contrary to August Weismann's Germ Plasm Theory and has led to several controversies. Recent data from metazoans as diverse as planarians, annelids and sea urchins reveal the presence of pluripotent stem cell populations that express germ plasm components, despite being considered to be somatic. These data also show that germ plasm is continuous in some of these animals, despite their discontinuous germline.
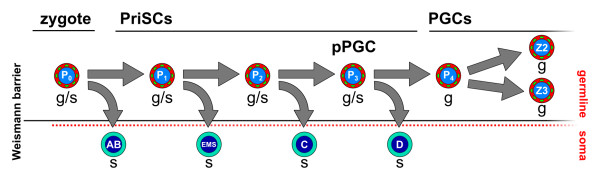
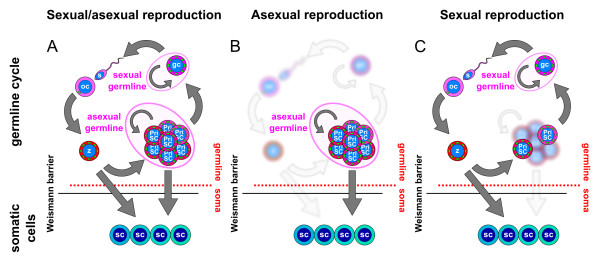
Presentation of the hypothesis: Here, based on recent molecular data on germ plasm components, I revise the germline concept. I introduce the concept of primordial stem cells, which are evolutionarily conserved stem cells that carry germ plasm components from the zygote to the germ cells. These cells, delineated by the classic concept of the Weismann barrier, can contribute to different extents to somatic tissues or be present in a rudimentary state. The primordial stem cells are a part of the germline that can drive asexual reproduction.
Testing the hypothesis: Molecular information on the expression of germ plasm components is needed during early development of non-classic model organisms, with special attention to those capable of undergoing asexual reproduction and regeneration. The cell lineage of germ plasm component-containing cells will also shed light on their position with respect to the Weismann barrier. This information will help in understanding the germline and its associated stem cells across metazoan phylogeny.
Implications of the hypothesis: This revision of the germline concept explains the extensive similarities observed among stem cells and germline cells in a wide variety of animals, and predicts the expression of germ plasm components in many others. The life history of these animals can be simply explained by changes in the extent of self-renewal, proliferation and developmental potential of the primordial stem cells. The inclusion of the primordial stem cells as a part of the germline, therefore, solves many controversies and provides a continuous germline, just as originally envisaged by August Weismann.
Figures



References
-
- Weismann A. Das Keimplasma: eine Theorie der Vererbung. Jena: Fischer; 1892.
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
