

Cell cycle stage-specific roles of Rad18 in tolerance and repair of oxidative DNA damage
- PMID: 23295675
- PMCID: PMC3575850
- DOI: 10.1093/nar/gks1325
Cell cycle stage-specific roles of Rad18 in tolerance and repair of oxidative DNA damage
Abstract
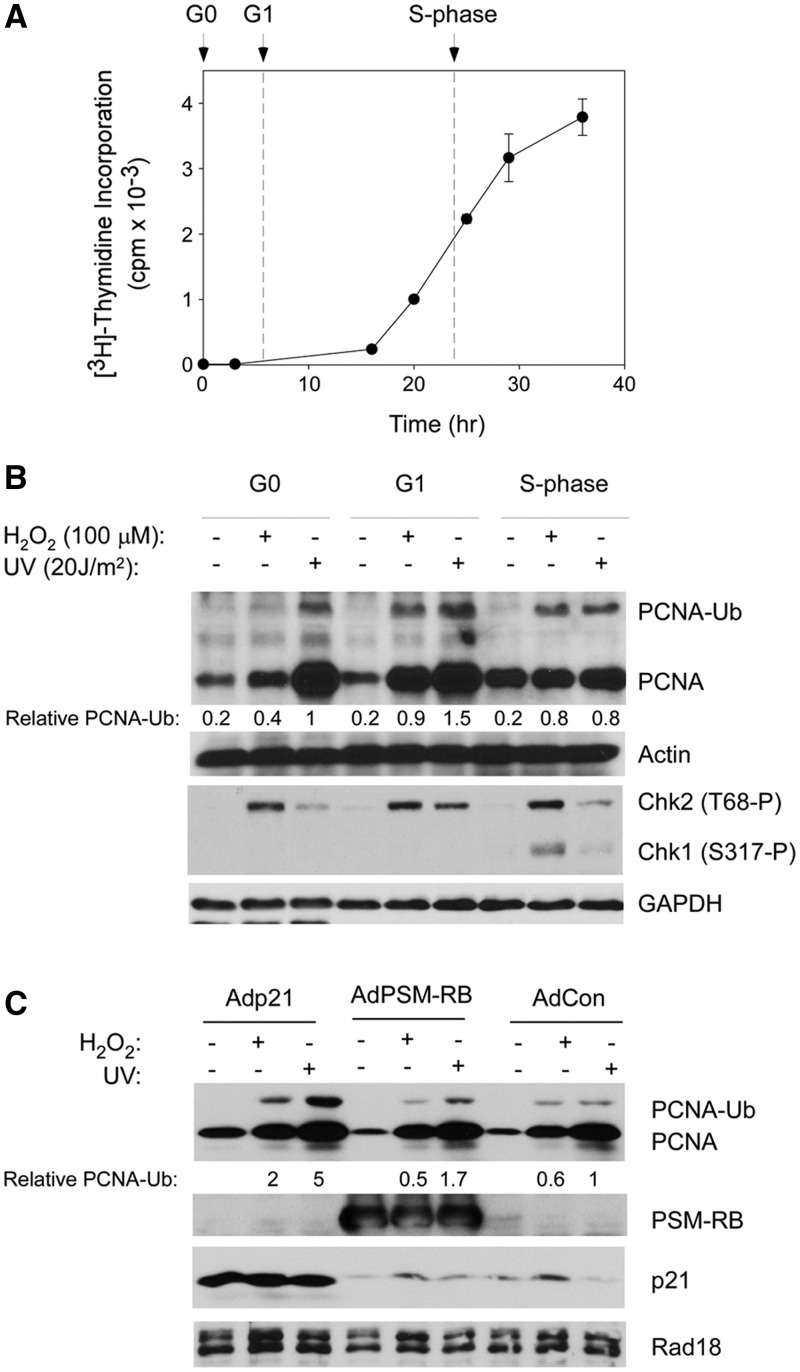
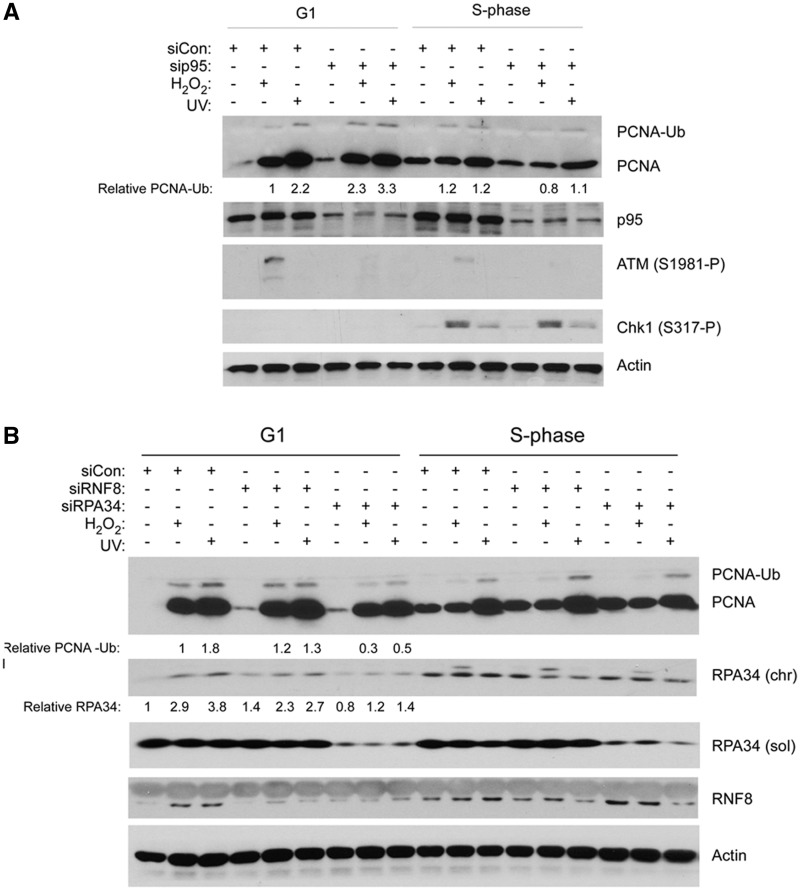
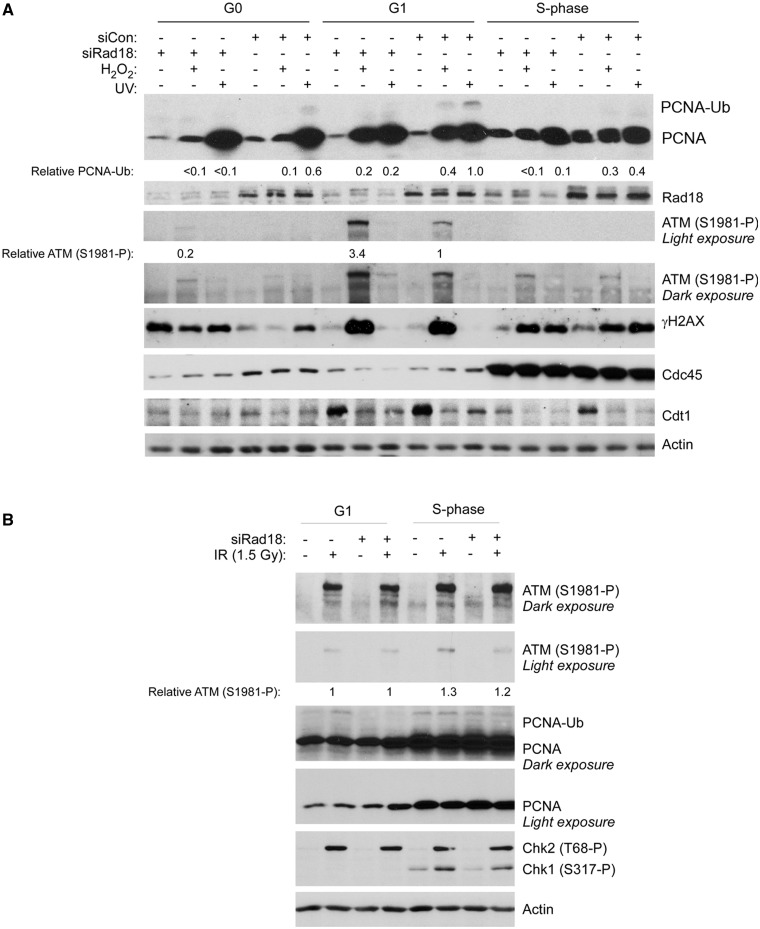
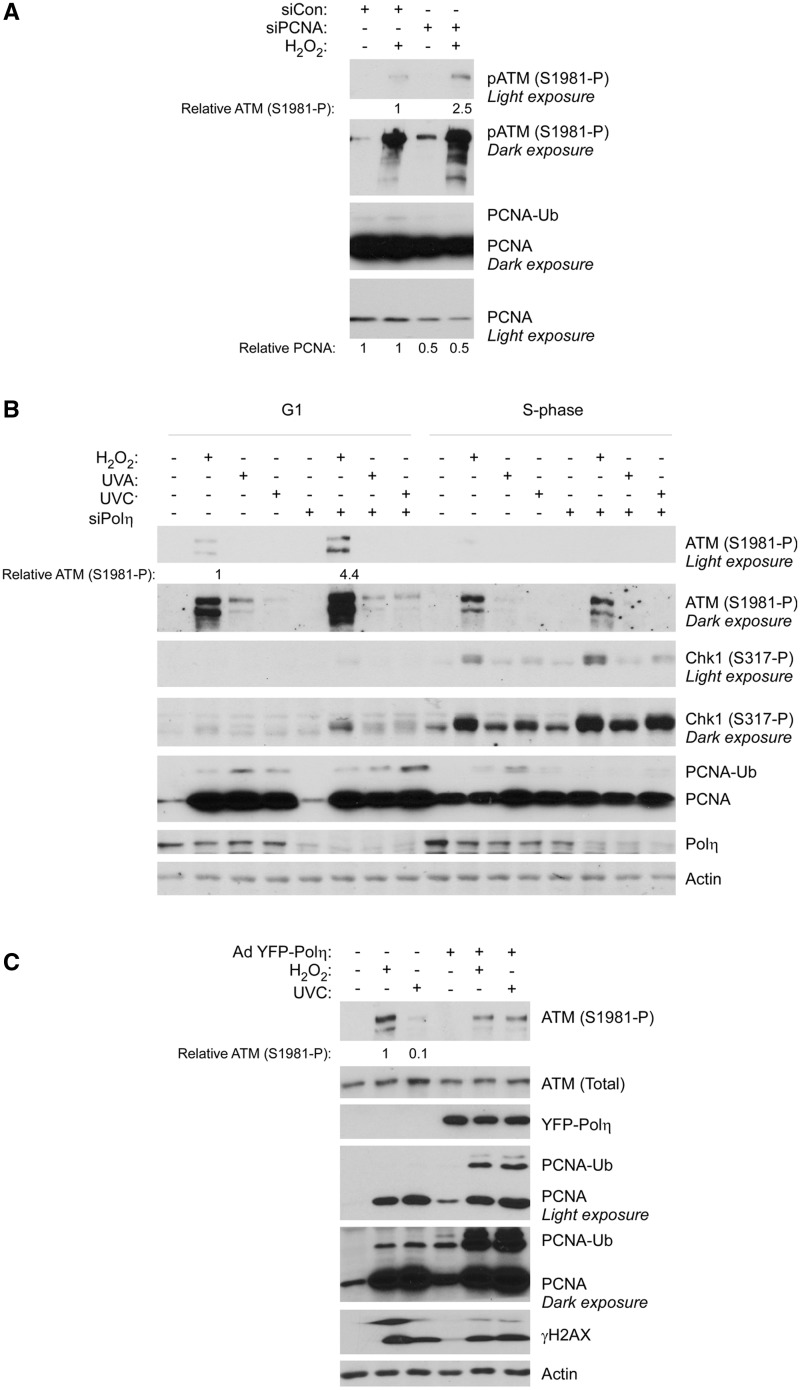
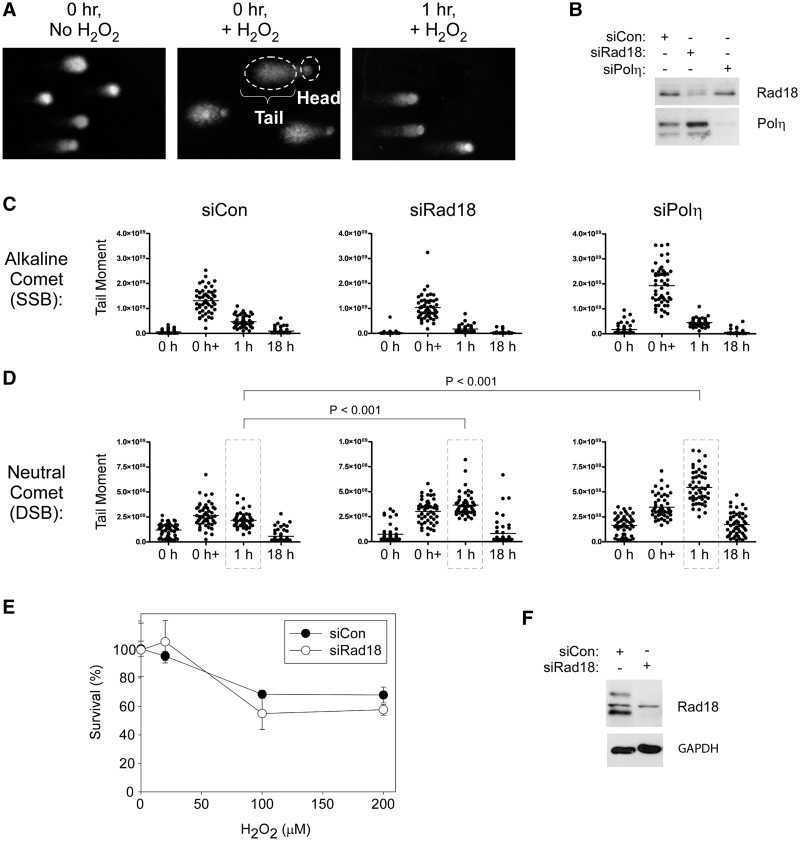
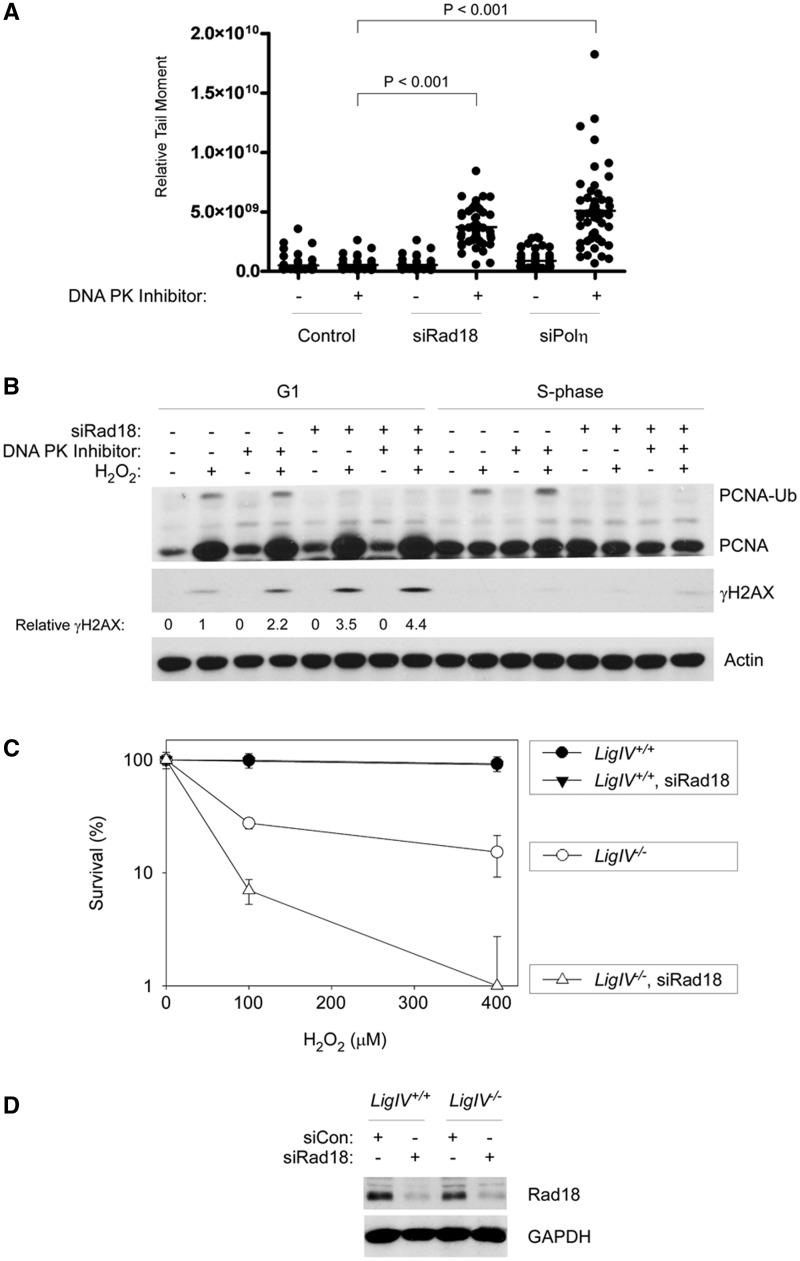
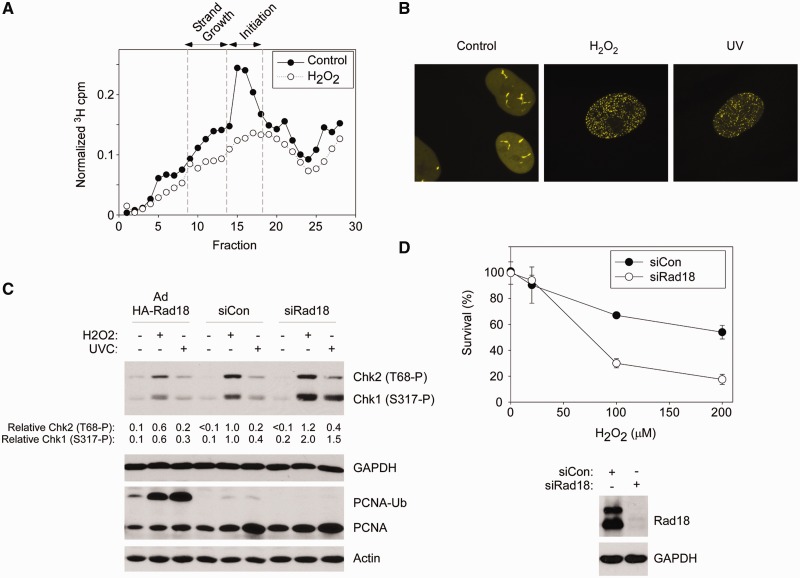
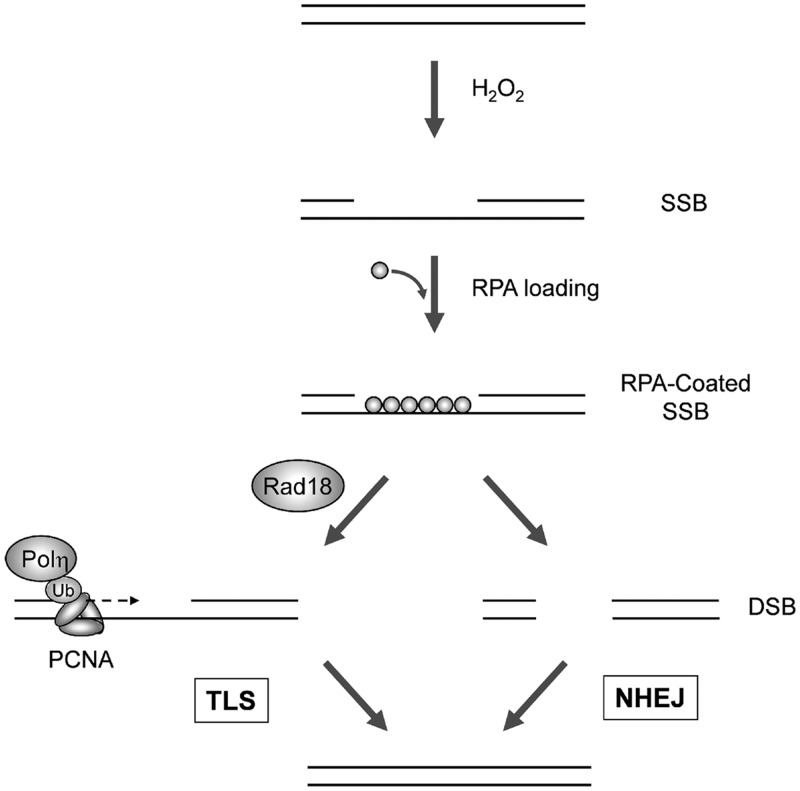
The E3 ubiquitin ligase Rad18 mediates tolerance of replication fork-stalling bulky DNA lesions, but whether Rad18 mediates tolerance of bulky DNA lesions acquired outside S-phase is unclear. Using synchronized cultures of primary human cells, we defined cell cycle stage-specific contributions of Rad18 to genome maintenance in response to ultraviolet C (UVC) and H(2)O(2)-induced DNA damage. UVC and H(2)O(2) treatments both induced Rad18-mediated proliferating cell nuclear antigen mono-ubiquitination during G(0), G(1) and S-phase. Rad18 was important for repressing H(2)O(2)-induced (but not ultraviolet-induced) double strand break (DSB) accumulation and ATM S1981 phosphorylation only during G(1), indicating a specific role for Rad18 in processing of oxidative DNA lesions outside S-phase. However, H(2)O(2)-induced DSB formation in Rad18-depleted G1 cells was not associated with increased genotoxin sensitivity, indicating that back-up DSB repair mechanisms compensate for Rad18 deficiency. Indeed, in DNA LigIV-deficient cells Rad18-depletion conferred H(2)O(2)-sensitivity, demonstrating functional redundancy between Rad18 and non-homologous end joining for tolerance of oxidative DNA damage acquired during G(1). In contrast with G(1)-synchronized cultures, S-phase cells were H(2)O(2)-sensitive following Rad18-depletion. We conclude that although Rad18 pathway activation by oxidative lesions is not restricted to S-phase, Rad18-mediated trans-lesion synthesis by Polη is dispensable for damage-tolerance in G(1) (because of back-up non-homologous end joining-mediated DSB repair), yet Rad18 is necessary for damage tolerance during S-phase.
Figures
References
-
- Prakash S, Johnson RE, Prakash L. Eukaryotic translesion synthesis DNA polymerases: specificity of structure and function. Annu. Rev. Biochem. 2005;74:317–353. - PubMed
-
- Ohmori H, Friedberg EC, Fuchs RP, Goodman MF, Hanaoka F, Hinkle D, Kunkel TA, Lawrence CW, Livneh Z, Nohmi T, et al. The Y-family of DNA polymerases. Mol. Cell. 2001;8:7–8. - PubMed
-
- Johnson RE, Prakash S, Prakash L. Efficient bypass of a thymine-thymine dimer by yeast DNA polymerase, Poleta. Science. 1999;283:1001–1004. - PubMed
-
- Masutani C, Kusumoto R, Yamada A, Dohmae N, Yokoi M, Yuasa M, Araki M, Iwai S, Takio K, Hanaoka F. The XPV (xeroderma pigmentosum variant) gene encodes human DNA polymerase eta. Nature. 1999;399:700–704. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
