

Cell death atlas of the postnatal mouse ventral forebrain and hypothalamus: effects of age and sex
- PMID: 23296992
- PMCID: PMC4968939
- DOI: 10.1002/cne.23298
Cell death atlas of the postnatal mouse ventral forebrain and hypothalamus: effects of age and sex
Abstract
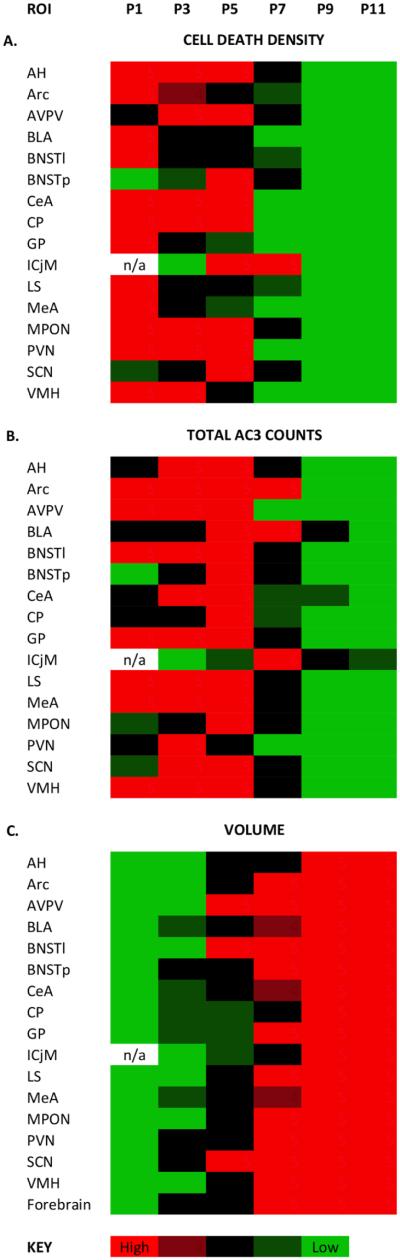
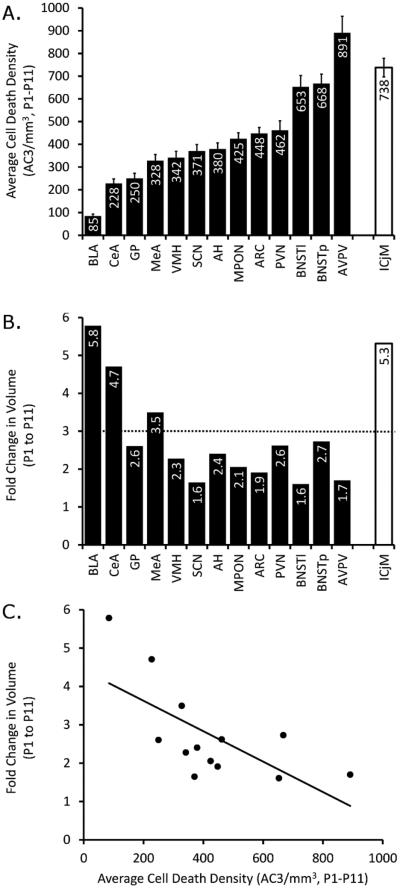
Naturally occurring cell death is essential to the development of the mammalian nervous system. Although the importance of developmental cell death has been appreciated for decades, there is no comprehensive account of cell death across brain areas in the mouse. Moreover, several regional sex differences in cell death have been described for the ventral forebrain and hypothalamus, but it is not known how widespread the phenomenon is. We used immunohistochemical detection of activated caspase-3 to identify dying cells in the brains of male and female mice from postnatal day (P) 1 to P11. Cell death density, total number of dying cells, and regional volume were determined in 16 regions of the hypothalamus and ventral forebrain (the anterior hypothalamus, arcuate nucleus, anteroventral periventricular nucleus, medial preoptic nucleus, paraventricular nucleus, suprachiasmatic nucleus, and ventromedial nucleus of the hypothalamus; the basolateral, central, and medial amygdala; the lateral and principal nuclei of the bed nuclei of the stria terminalis; the caudate-putamen; the globus pallidus; the lateral septum; and the islands of Calleja). All regions showed a significant effect of age on cell death. The timing of peak cell death varied between P1 to P7, and the average rate of cell death varied tenfold among regions. Several significant sex differences in cell death and/or regional volume were detected. These data address large gaps in the developmental literature and suggest interesting region-specific differences in the prevalence and timing of cell death in the hypothalamus and ventral forebrain.
Copyright © 2013 Wiley Periodicals, Inc.
Figures
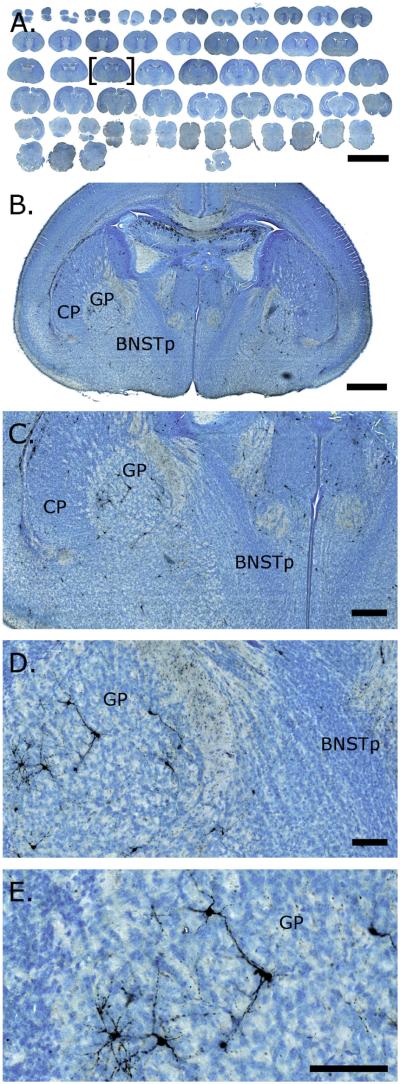

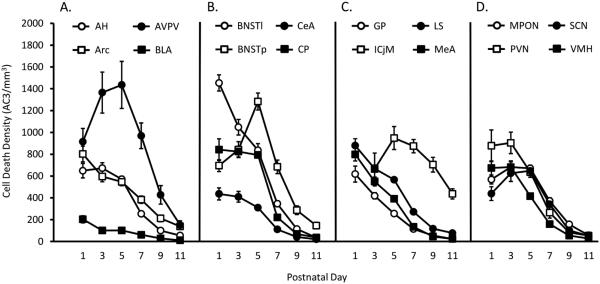
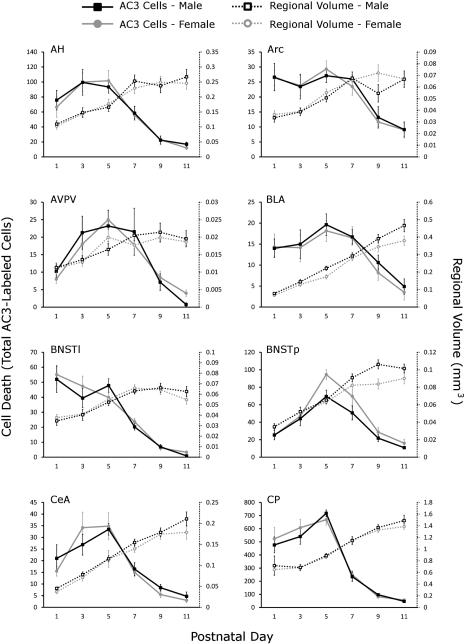
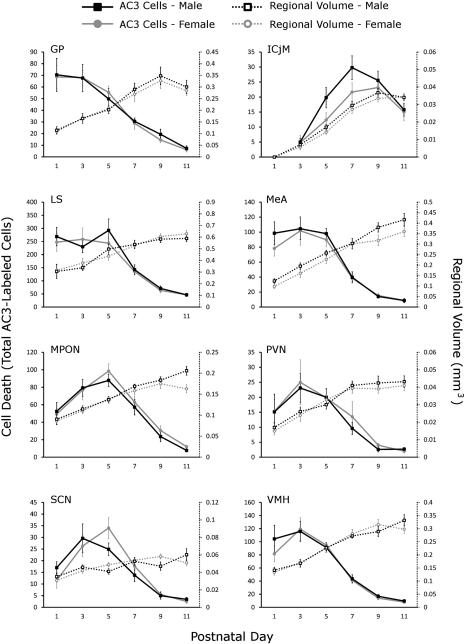

References
-
- Bayer SA, Altman J. Development of the preoptic area: time and site of origin, migratory routes, and settling patterns of its neurons. J Comp Neurol. 1987;265:65–95. - PubMed
-
- Berdel B, Moryś J, Maciejewska B, Dziewiatkowski J. Volume and topographical changes of the basolateral complex during the development of the rat’s amygdaloid body. Fol Morphol. 1997;56:1–11. - PubMed
-
- Blaschke AJ, Staley K, Chun J. Widespread programmed cell death in proliferative and postmitotic regions of the fetal cerebral cortex. Development. 1996;122:1165–1174. - PubMed
-
- Blaschke AJ, Weiner JA, Chun J. Programmed cell death is a universal feature of embryonic and postnatal neuroproliferative regions throughout the central nervous system. J Comp Neurol. 1998;396:39–50. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
