

Modulating the innate immune response by combinatorial engineering of endotoxin
- PMID: 23297218
- PMCID: PMC3557076
- DOI: 10.1073/pnas.1218080110
Modulating the innate immune response by combinatorial engineering of endotoxin
Abstract
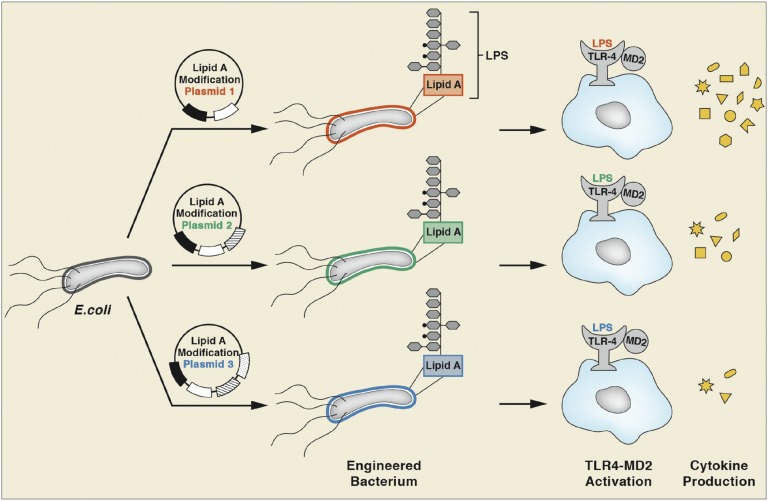
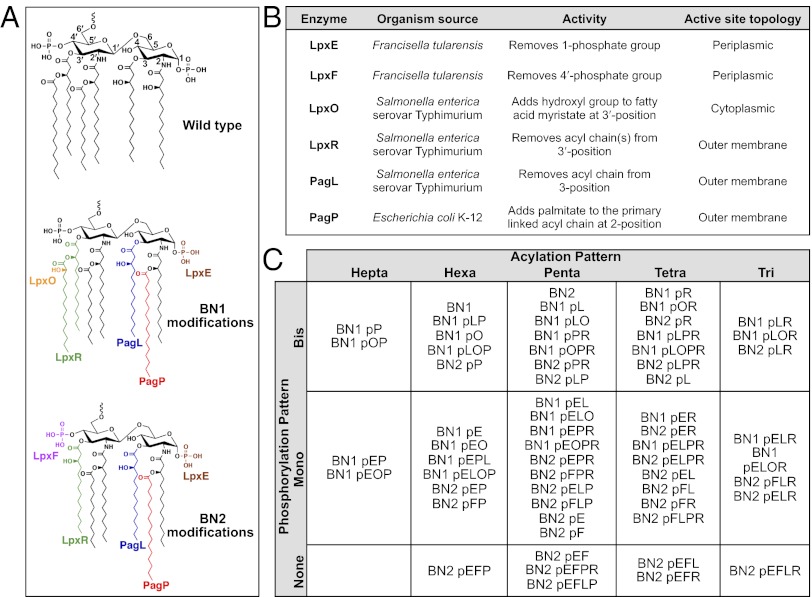
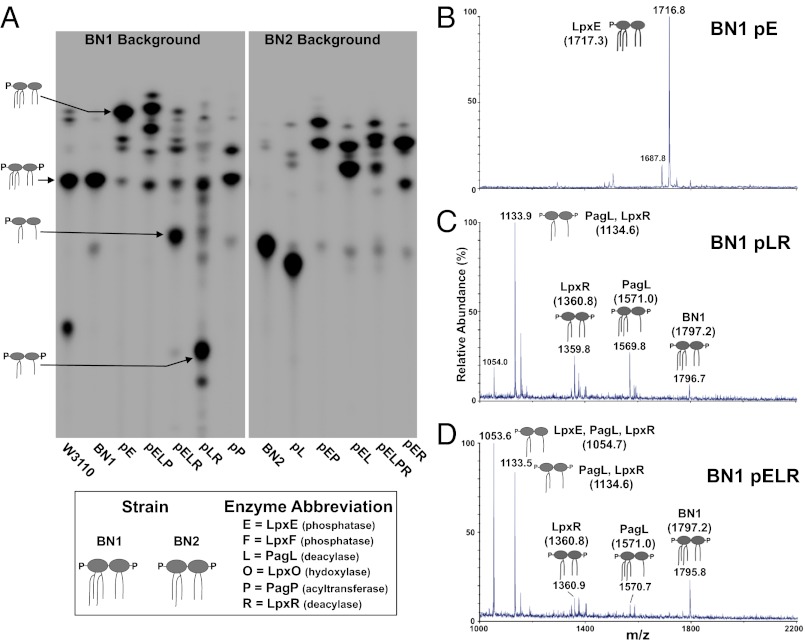
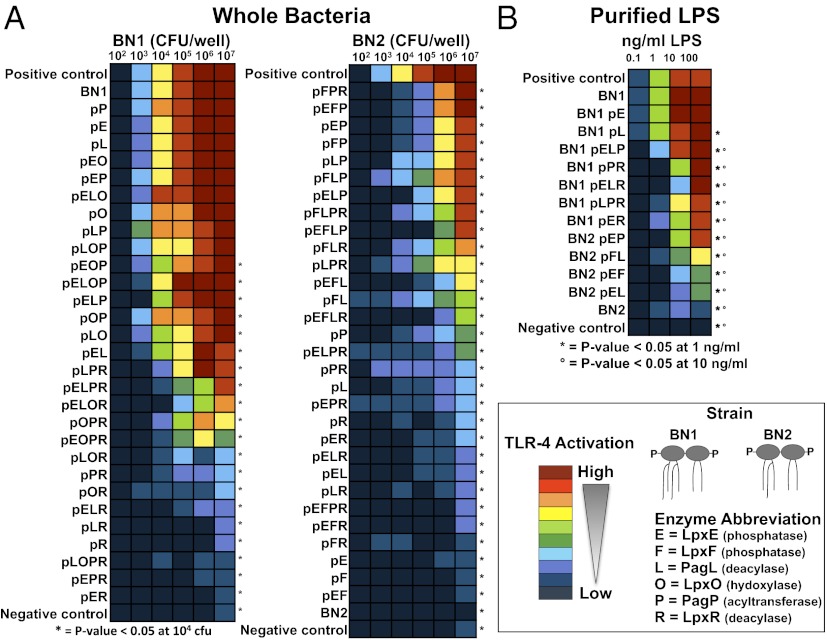
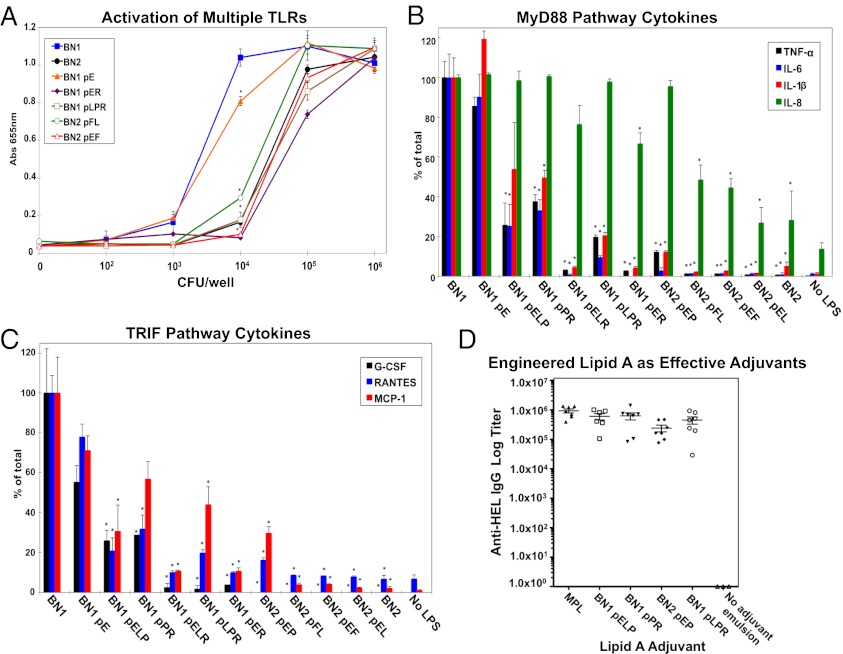
Despite its highly inflammatory nature, LPS is a molecule with remarkable therapeutic potential. Lipid A is a glycolipid that serves as the hydrophobic anchor of LPS and constitutes a potent ligand of the Toll-like receptor (TLR)4/myeloid differentiation factor 2 receptor of the innate immune system. A less toxic mixture of monophosphorylated lipid A species (MPL) recently became the first new Food and Drug Administration-approved adjuvant in over 70 y. Whereas wild-type Escherichia coli LPS provokes strong inflammatory MyD88 (myeloid differentiation primary response gene 88)-mediated TLR4 signaling, MPL preferentially induces less inflammatory TRIF (TIR-domain-containing adaptor-inducing IFN-β)-mediated responses. Here, we developed a system for combinatorial structural diversification of E. coli lipid A, yielding a spectrum of bioactive variants that display distinct TLR4 agonist activities and cytokine induction. Mice immunized with engineered lipid A/antigen emulsions exhibited robust IgG titers, indicating the efficacy of these molecules as adjuvants. This approach demonstrates how combinatorial engineering of lipid A can be exploited to generate a spectrum of immunostimulatory molecules for vaccine and therapeutics development.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Rationally Designed TLR4 Ligands for Vaccine Adjuvant Discovery.mBio. 2017 May 9;8(3):e00492-17. doi: 10.1128/mBio.00492-17. mBio. 2017. PMID: 28487429 Free PMC article.
-
The vaccine adjuvant monophosphoryl lipid A as a TRIF-biased agonist of TLR4.Science. 2007 Jun 15;316(5831):1628-32. doi: 10.1126/science.1138963. Science. 2007. PMID: 17569868
-
Adjuvant activity of naturally occurring monophosphoryl lipopolysaccharide preparations from mucosa-associated bacteria.Infect Immun. 2013 Sep;81(9):3317-25. doi: 10.1128/IAI.01150-12. Epub 2013 Jun 24. Infect Immun. 2013. PMID: 23798540 Free PMC article.
-
Chemistry of lipid A: at the heart of innate immunity.Chemistry. 2015 Jan 7;21(2):500-19. doi: 10.1002/chem.201403923. Epub 2014 Oct 29. Chemistry. 2015. PMID: 25353096 Review.
-
Taking a Toll on human disease: Toll-like receptor 4 agonists as vaccine adjuvants and monotherapeutic agents.Expert Opin Biol Ther. 2004 Jul;4(7):1129-38. doi: 10.1517/14712598.4.7.1129. Expert Opin Biol Ther. 2004. PMID: 15268679 Review.
Cited by
-
A Light-Controlled TLR4 Agonist and Selectable Activation of Cell Subpopulations.Chembiochem. 2015 Aug 17;16(12):1744-8. doi: 10.1002/cbic.201500164. Epub 2015 Jun 26. Chembiochem. 2015. PMID: 26097006 Free PMC article.
-
A Sensitive GC-MS Method for Quantitation of Lipid A Backbone Components and Terminal Phosphate Modifications.J Am Soc Mass Spectrom. 2022 Dec 7;33(12):2301-2309. doi: 10.1021/jasms.2c00266. Epub 2022 Nov 3. J Am Soc Mass Spectrom. 2022. PMID: 36326685 Free PMC article.
-
Convergence of Nanotechnology and Bacteriotherapy for Biomedical Applications.Adv Sci (Weinh). 2024 Apr;11(16):e2309295. doi: 10.1002/advs.202309295. Epub 2024 Feb 15. Adv Sci (Weinh). 2024. PMID: 38358998 Free PMC article. Review.
-
A cell-free biosynthesis platform for modular construction of protein glycosylation pathways.Nat Commun. 2019 Nov 27;10(1):5404. doi: 10.1038/s41467-019-12024-9. Nat Commun. 2019. PMID: 31776339 Free PMC article.
-
Control freaks-signals and cues governing the regulation of virulence in attaching and effacing pathogens.Biochem Soc Trans. 2019 Feb 28;47(1):229-238. doi: 10.1042/BST20180546. Epub 2018 Dec 17. Biochem Soc Trans. 2019. PMID: 30559275 Free PMC article. Review.
References
-
- Pfeiffer R. Untersuchungen uber das Choleragift. Z Hygeine. 1892;11:393–412. German.
-
- Park BS, et al. The structural basis of lipopolysaccharide recognition by the TLR4-MD-2 complex. Nature. 2009;458(7242):1191–1195. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
