

Nuclear export inhibition through covalent conjugation and hydrolysis of Leptomycin B by CRM1
- PMID: 23297231
- PMCID: PMC3557022
- DOI: 10.1073/pnas.1217203110
Nuclear export inhibition through covalent conjugation and hydrolysis of Leptomycin B by CRM1
Abstract
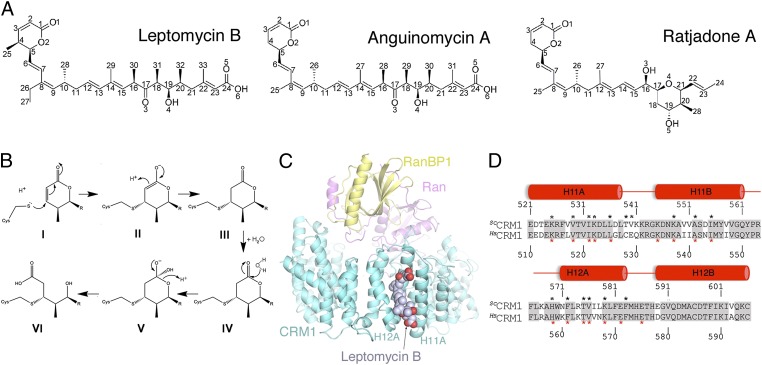
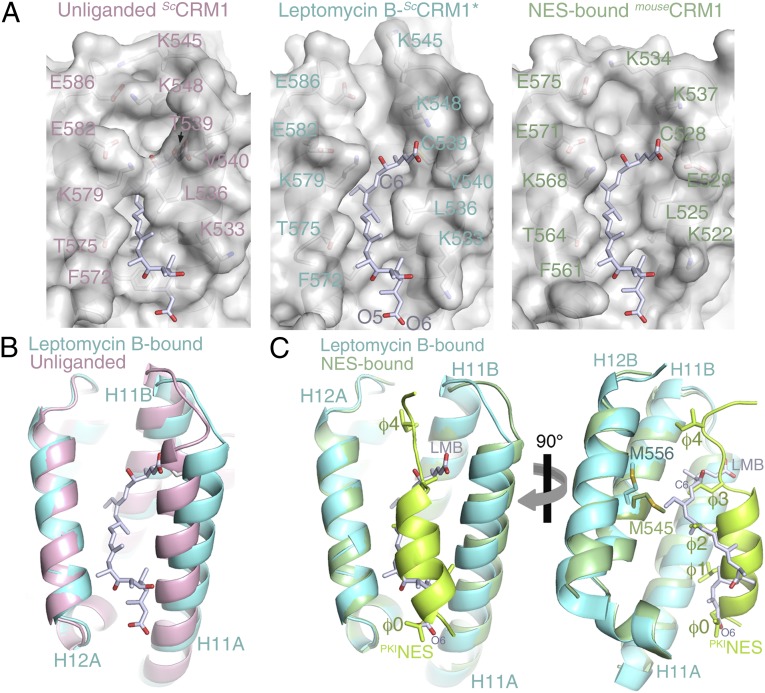
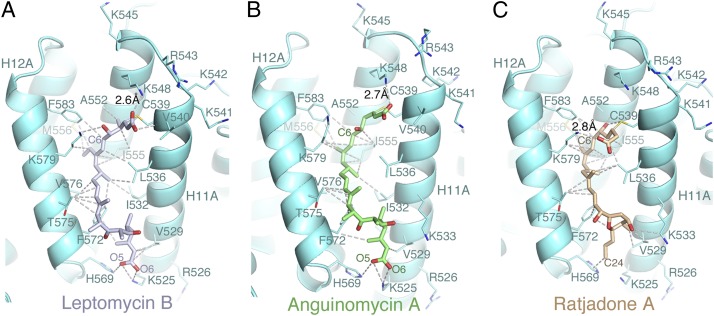
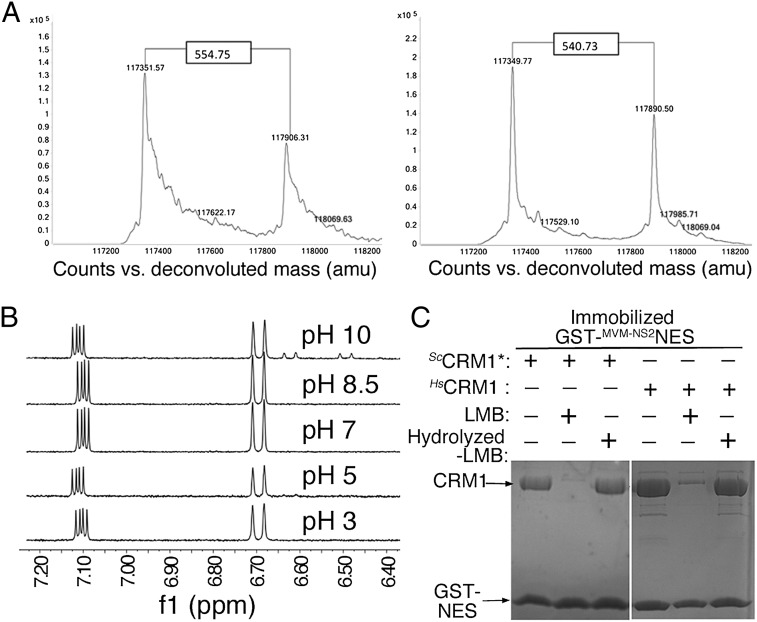
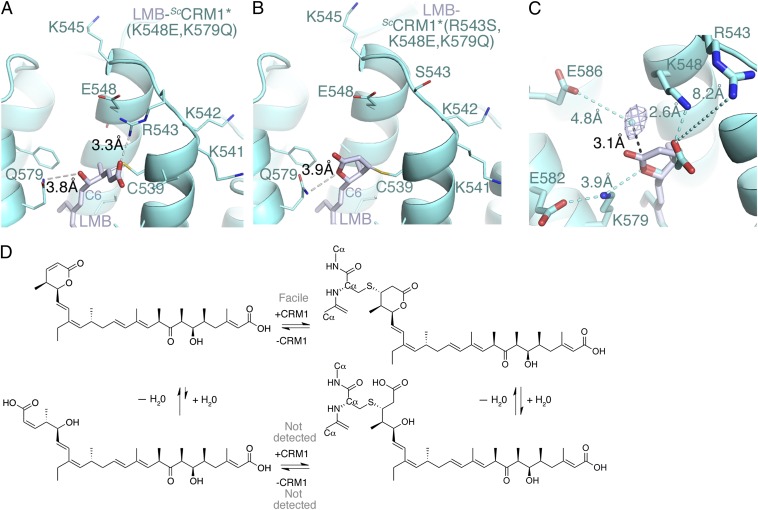
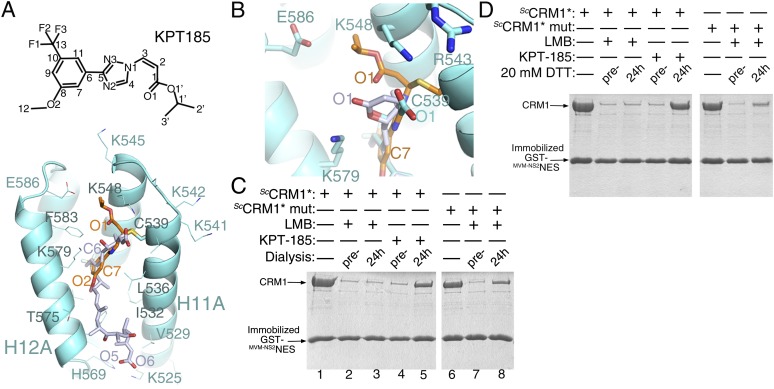
The polyketide natural product Leptomycin B inhibits nuclear export mediated by the karyopherin protein chromosomal region maintenance 1 (CRM1). Here, we present 1.8- to 2.0-Å-resolution crystal structures of CRM1 bound to Leptomycin B and related inhibitors Anguinomycin A and Ratjadone A. Structural and complementary chemical analyses reveal an unexpected mechanism of inhibition involving covalent conjugation and CRM1-mediated hydrolysis of the natural products' lactone rings. Furthermore, mutagenesis reveals the mechanism of hydrolysis by CRM1. The nuclear export signal (NES)-binding groove of CRM1 is able to drive a chemical reaction in addition to binding protein cargoes for transport through the nuclear pore complex.
Conflict of interest statement
Conflict of interest statement: Y.M.C. is a consultant for Karyopharm Therapeutics.
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
