

Nascent-Seq analysis of Drosophila cycling gene expression
- PMID: 23297234
- PMCID: PMC3557077
- DOI: 10.1073/pnas.1219969110
Nascent-Seq analysis of Drosophila cycling gene expression
Abstract
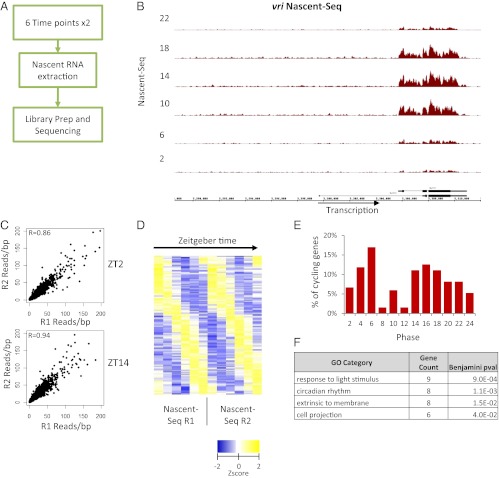
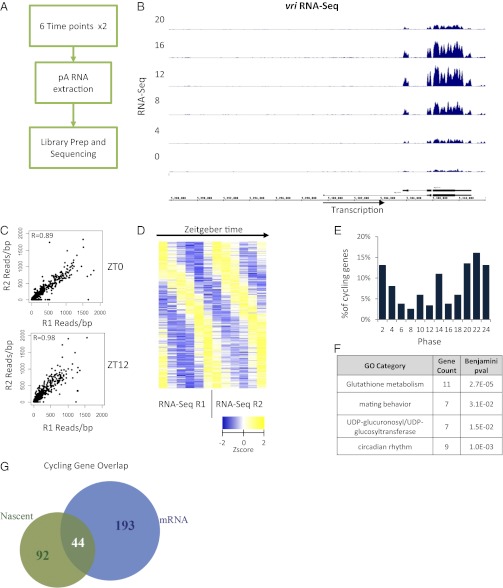
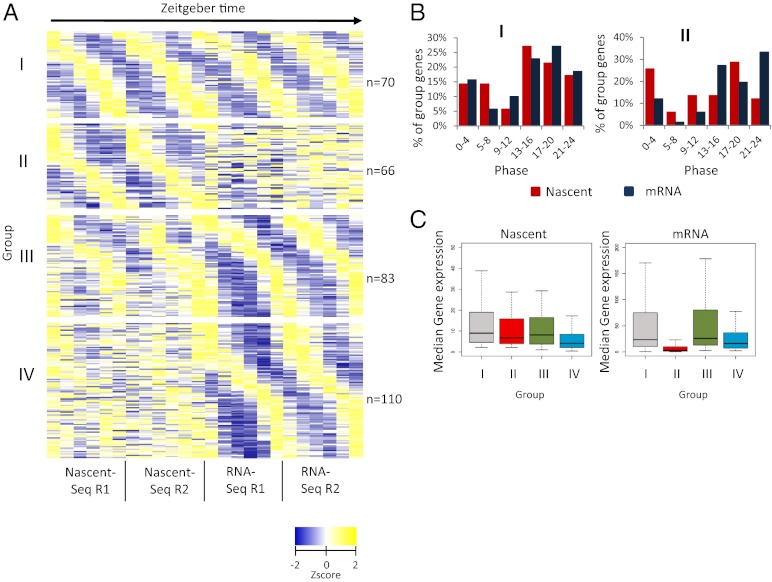
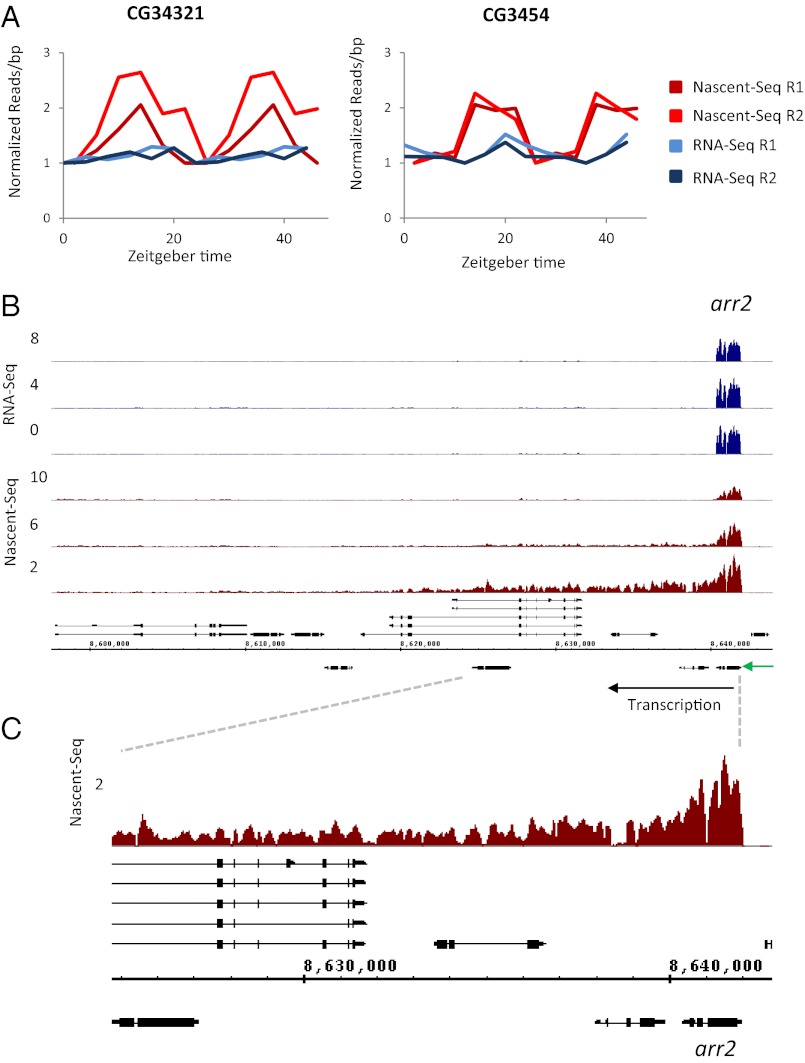
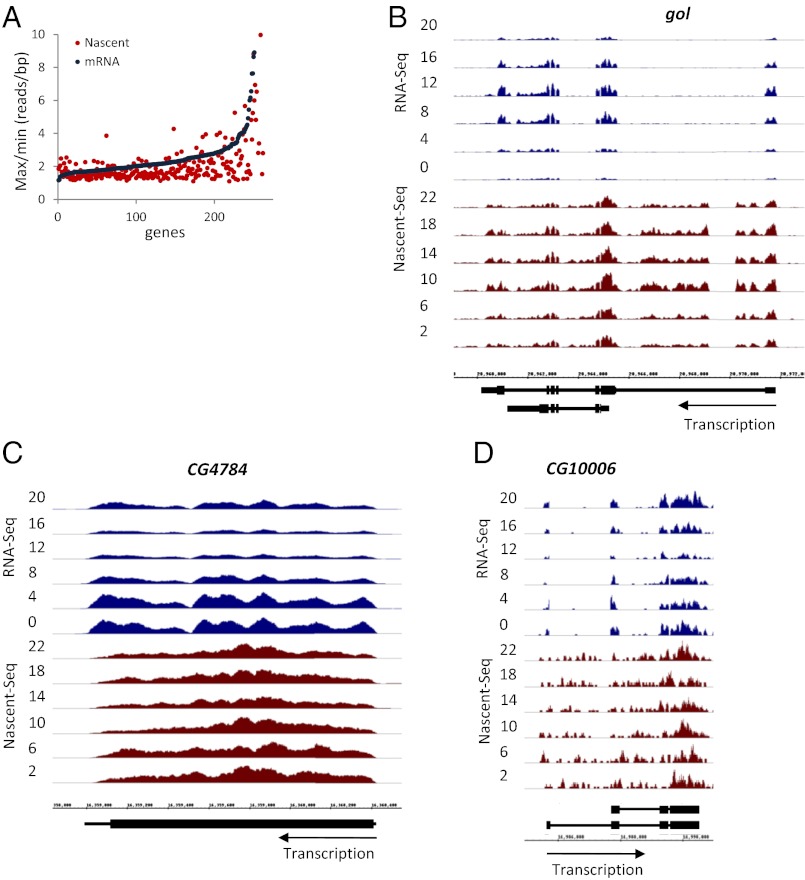
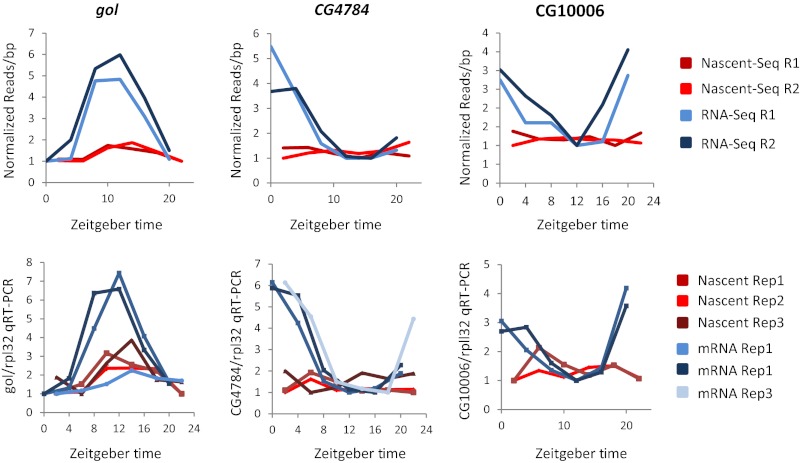
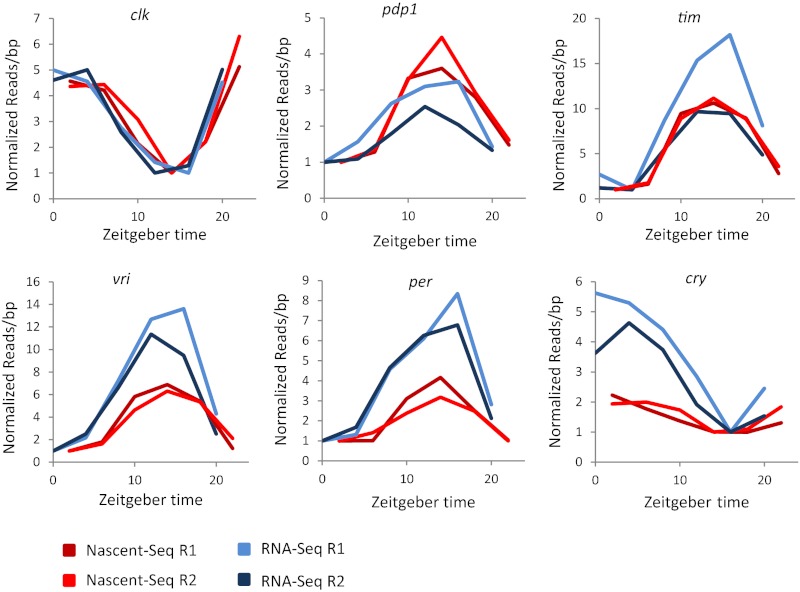
Rhythmic mRNA expression is a hallmark of circadian biology and has been described in numerous experimental systems including mammals. A small number of core clock gene mRNAs and a much larger number of output mRNAs are under circadian control. The rhythmic expression of core clock genes is regulated at the transcriptional level, and this regulation is important for the timekeeping mechanism. However, the relative contribution of transcriptional and post transcriptional regulation to global circadian mRNA oscillations is unknown. To address this issue in Drosophila, we isolated nascent RNA from adult fly heads collected at different time points and subjected it to high-throughput sequencing. mRNA was isolated and sequence din parallel. Some genes had cycling nascent RNAs with no cycling mRNA, caused,most likely, by light-mediated read-through transcription. Most genes with cycling mRNAs had significant nascent RNA cycling amplitudes, indicating a prominent role for circadian transcriptional regulation. However, a considerable fraction had higher mRNA amplitudes than nascent RNA amplitudes. The same comparison for core clock gene mRNAs gives rise to a qualitatively similar conclusion. The data therefore indicate a significant quantitative contribution of post transcriptional regulation to mRNA cycling.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
