

The soluble interleukin-6 receptor is a mediator of hematopoietic and skeletal actions of parathyroid hormone
- PMID: 23297399
- PMCID: PMC3591592
- DOI: 10.1074/jbc.M112.393363
The soluble interleukin-6 receptor is a mediator of hematopoietic and skeletal actions of parathyroid hormone
Abstract
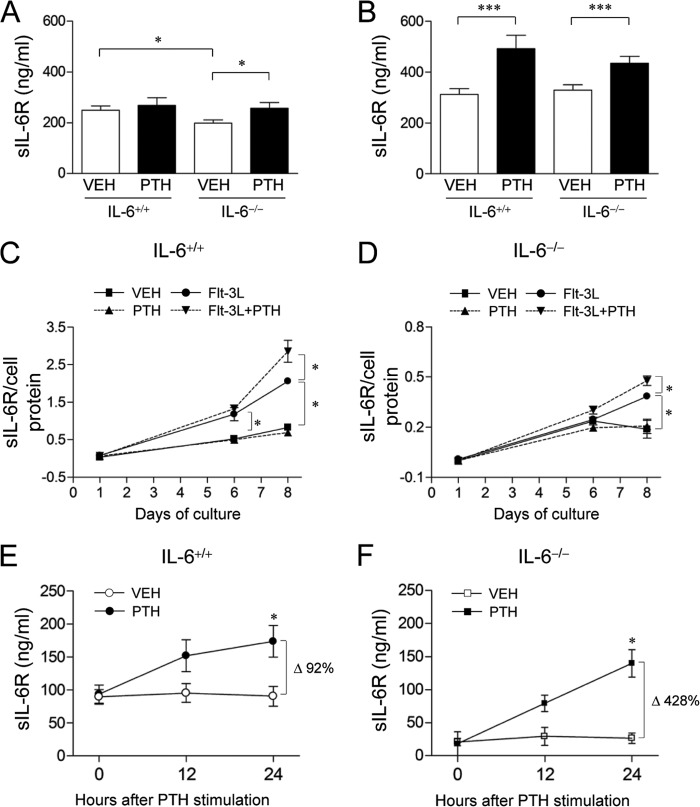
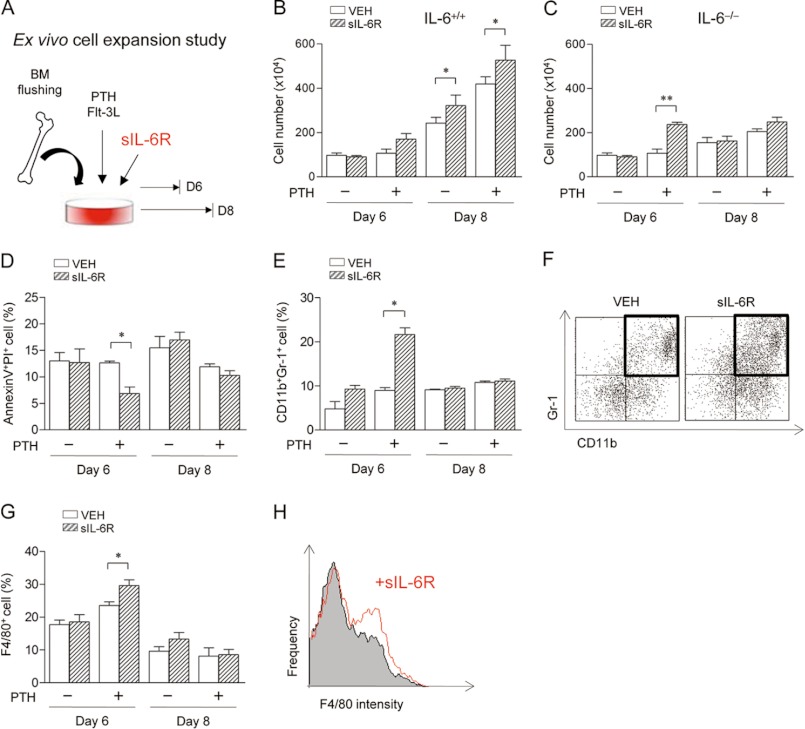
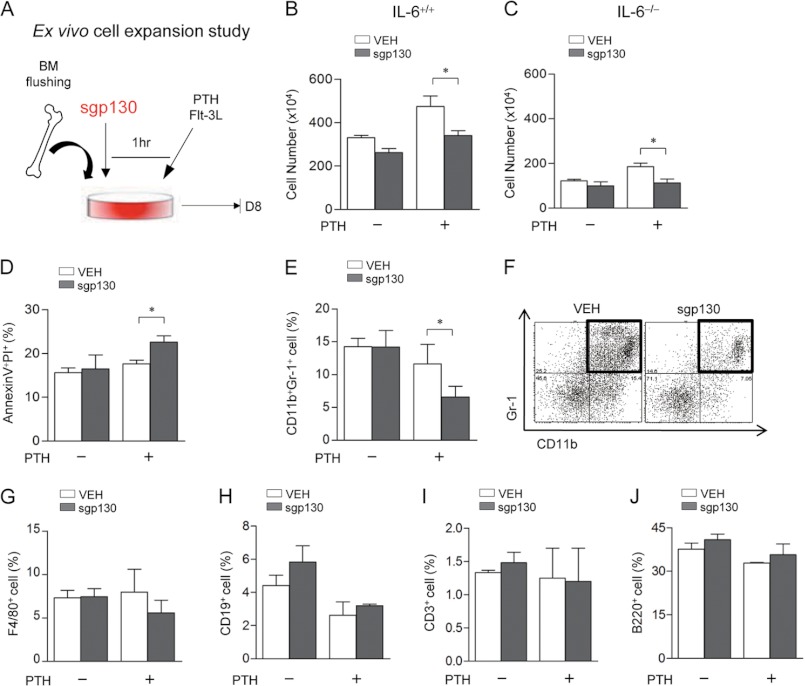
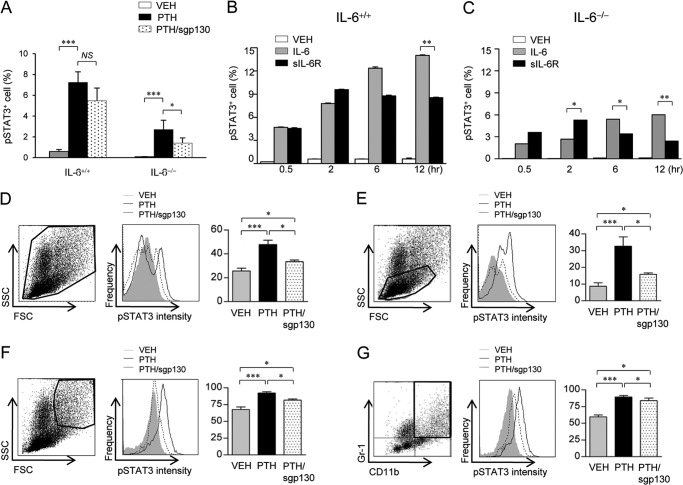
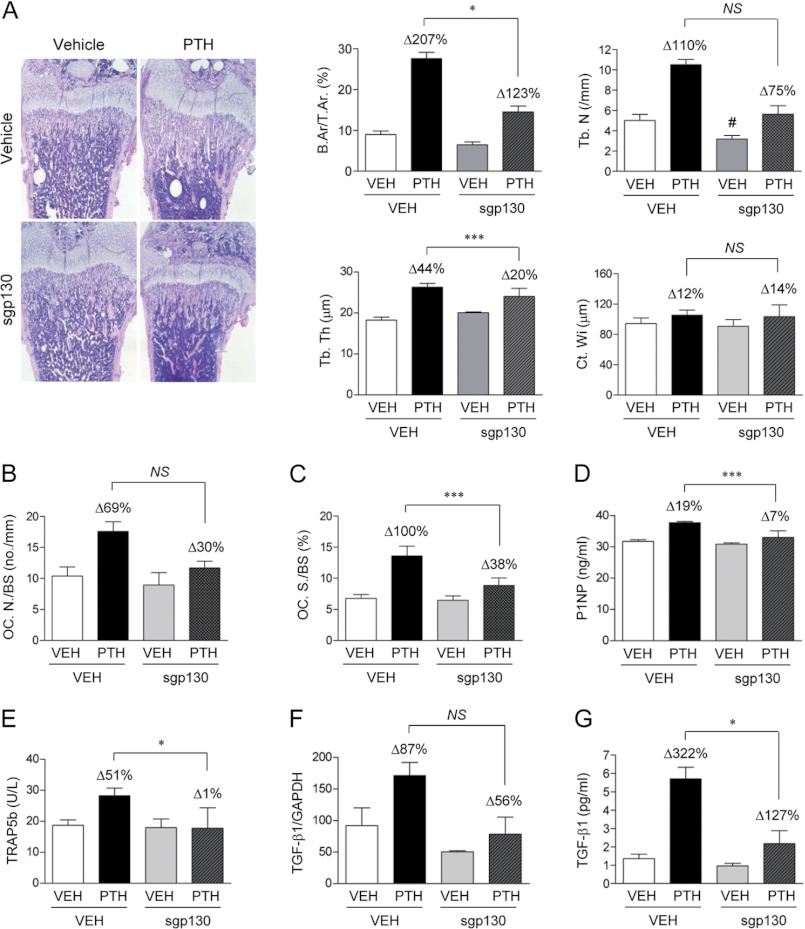
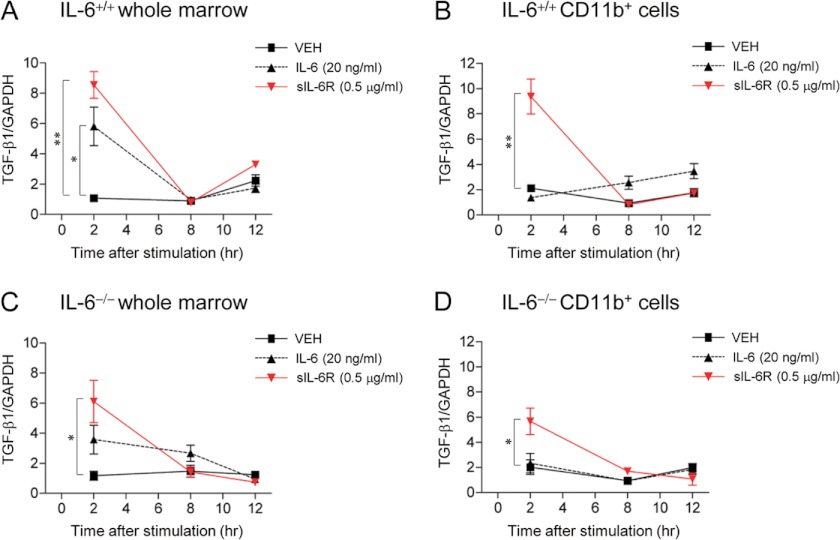
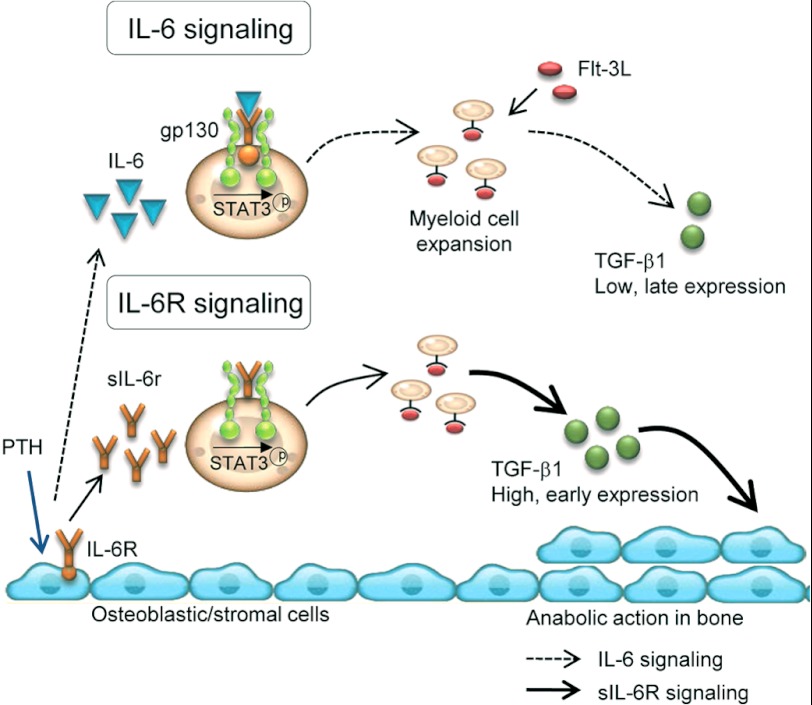
Both PTH and IL-6 signaling play pivotal roles in hematopoiesis and skeletal biology, but their interdependence is unclear. The purpose of this study was to evaluate the effect of IL-6 and soluble IL-6 receptor (sIL-6R) on hematopoietic and skeletal actions of PTH. In the bone microenvironment, PTH stimulated sIL-6R protein levels in primary osteoblast cultures in vitro and bone marrow in vivo in both IL-6(+/+) and IL-6(-/-) mice. PTH-mediated hematopoietic cell expansion was attenuated in IL-6(-/-) compared with IL-6(+/+) bone marrow, whereas sIL-6R treatment amplified PTH actions in IL-6(-/-) earlier than IL-6(+/+) marrow cultures. Blocking sIL-6R signaling with sgp130 (soluble glycoprotein 130 receptor) inhibited PTH-dependent hematopoietic cell expansion in IL-6(-/-) marrow. In the skeletal system, although intermittent PTH administration to IL-6(+/+) and IL-6(-/-) mice resulted in similar anabolic actions, blocking sIL-6R significantly attenuated PTH anabolic actions. sIL-6R showed no direct effects on osteoblast proliferation or differentiation in vitro; however, it up-regulated myeloid cell expansion and production of the mesenchymal stem cell recruiting agent, TGF-β1 in the bone marrow microenvironment. Collectively, sIL-6R demonstrated orphan function and mediated PTH anabolic actions in bone in association with support of myeloid lineage cells in the hematopoietic system.
Figures
References
-
- Demiralp B., Chen H. L., Koh A. J., Keller E. T., McCauley L. K. (2002) Anabolic actions of parathyroid hormone during bone growth are dependent on c-fos. Endocrinology 143, 4038–4047 - PubMed
-
- Chang J. K., Chang L. H., Hung S. H., Wu S. C., Lee H. Y., Lin Y. S., Chen C. H., Fu Y. C., Wang G. J., Ho M. L. (2009) Parathyroid hormone 1–34 inhibits terminal differentiation of human articular chondrocytes and osteoarthritis progression in rats. Arthritis Rheum. 60, 3049–3060 - PubMed
-
- Calvi L. M., Adams G. B., Weibrecht K. W., Weber J. M., Olson D. P., Knight M. C., Martin R. P., Schipani E., Divieti P., Bringhurst F. R., Milner L. A., Kronenberg H. M., Scadden D. T. (2003) Osteoblastic cells regulate the haematopoietic stem cell niche. Nature 425, 841–846 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
