

Scanning the topography of polyamine blocker binding in an inwardly rectifying potassium channel
- PMID: 23300089
- PMCID: PMC3585099
- DOI: 10.1074/jbc.M112.383794
Scanning the topography of polyamine blocker binding in an inwardly rectifying potassium channel
Abstract
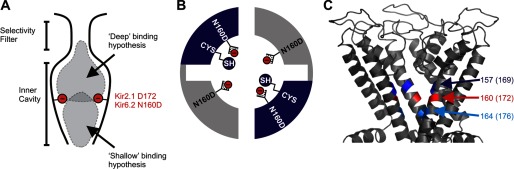
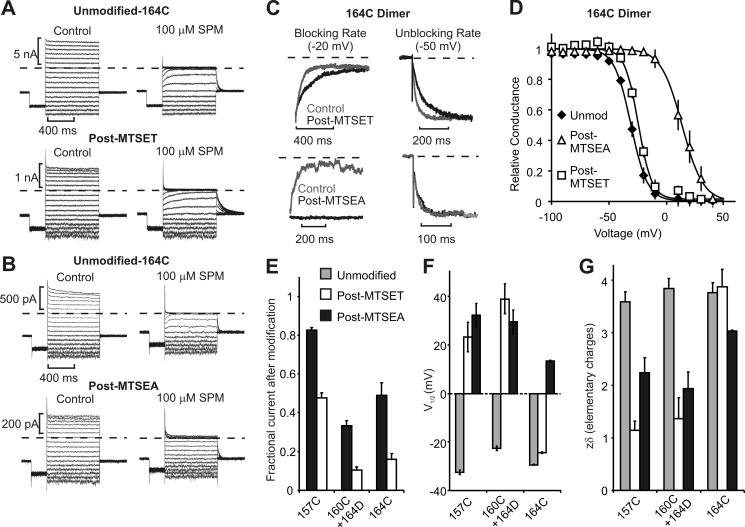
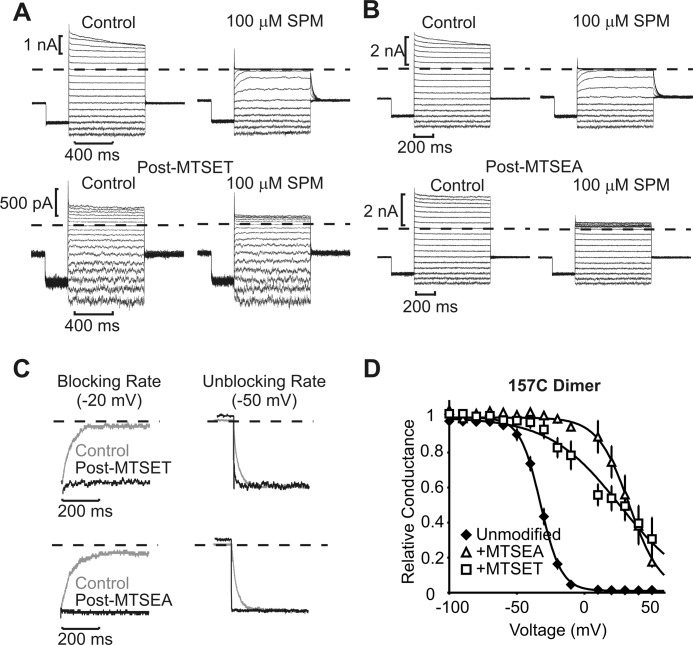
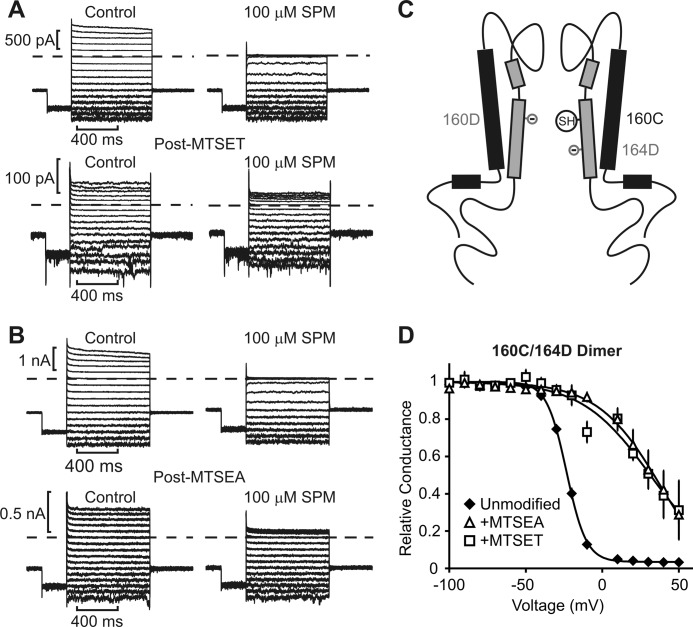
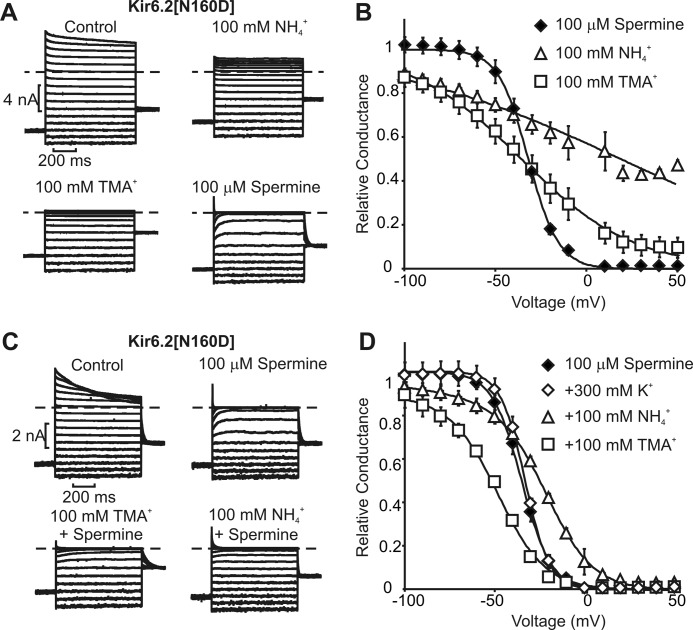
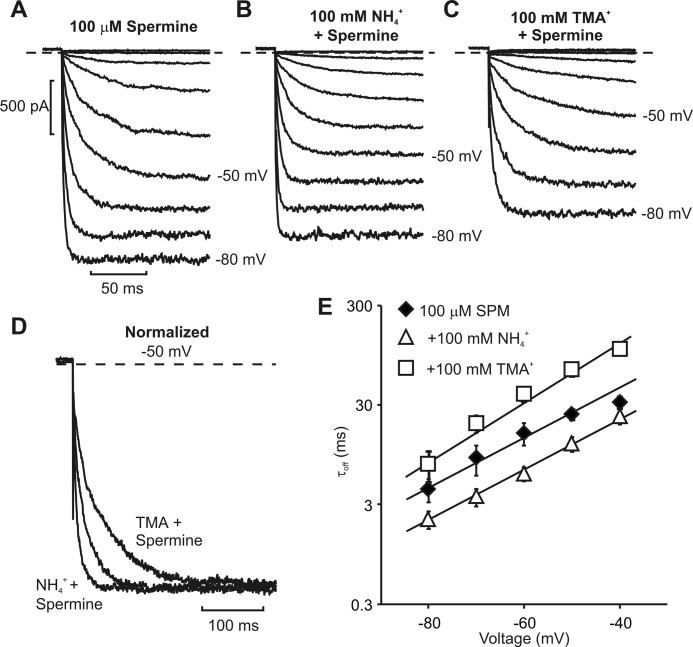
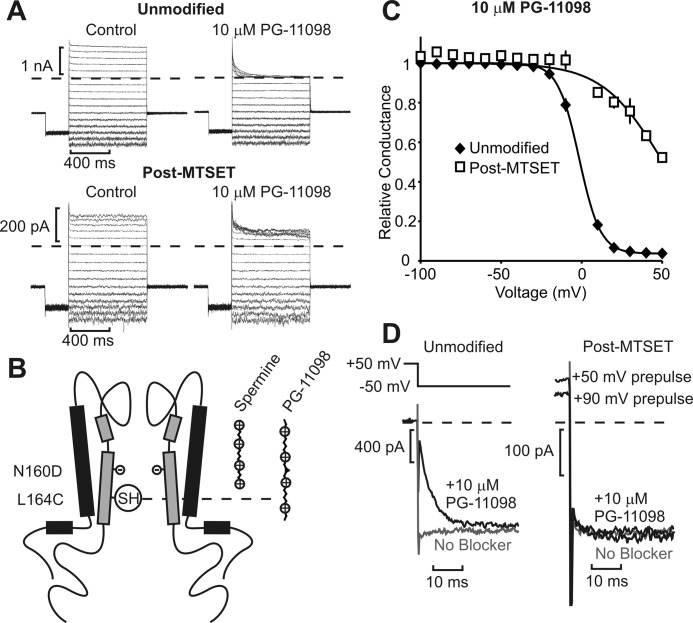
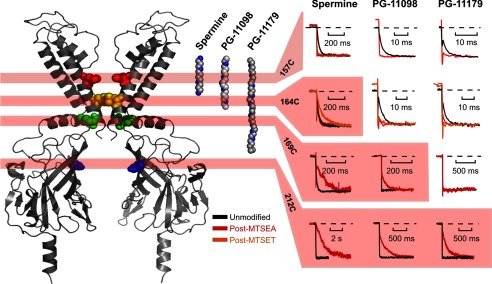
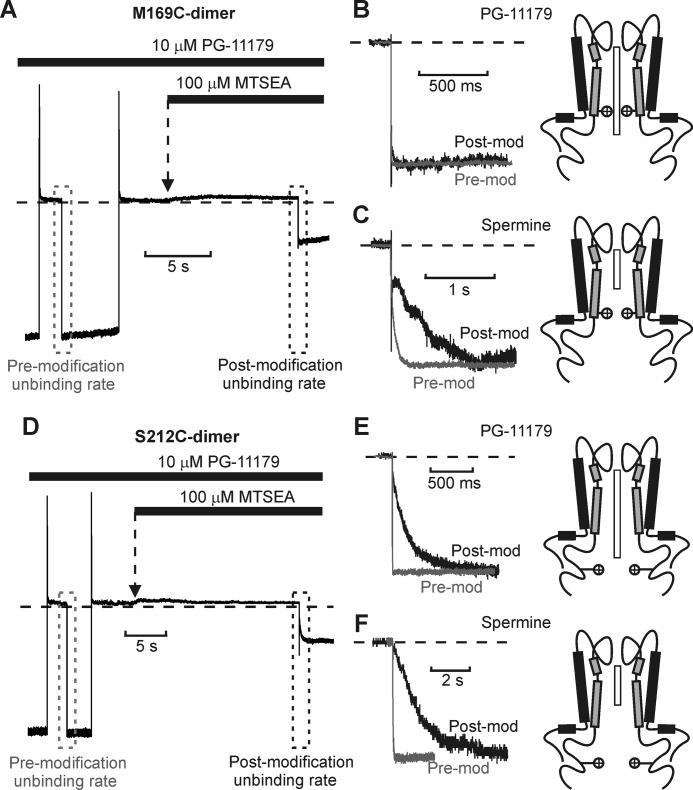
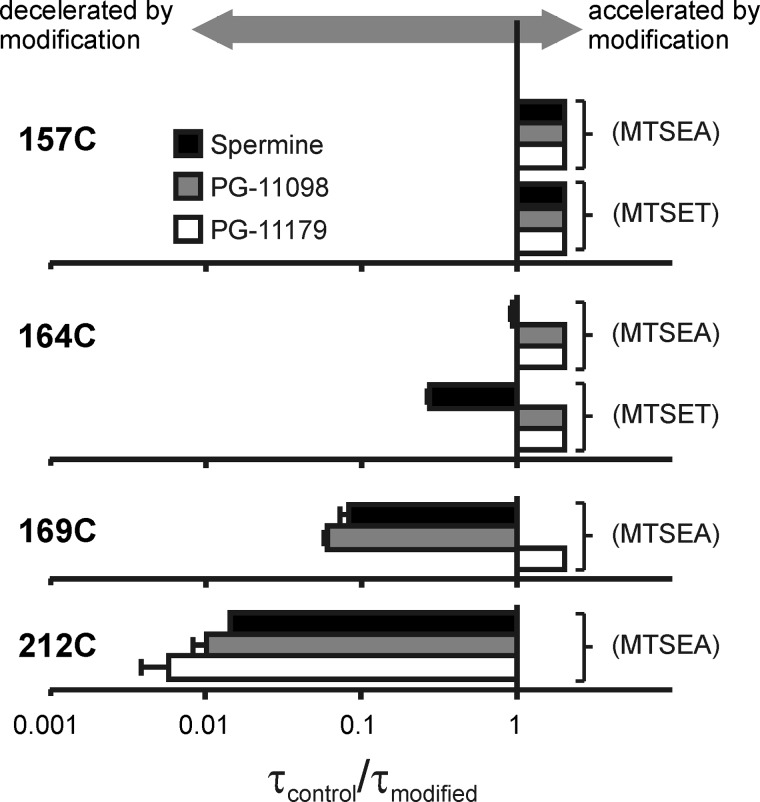
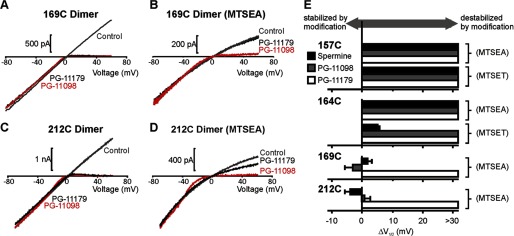
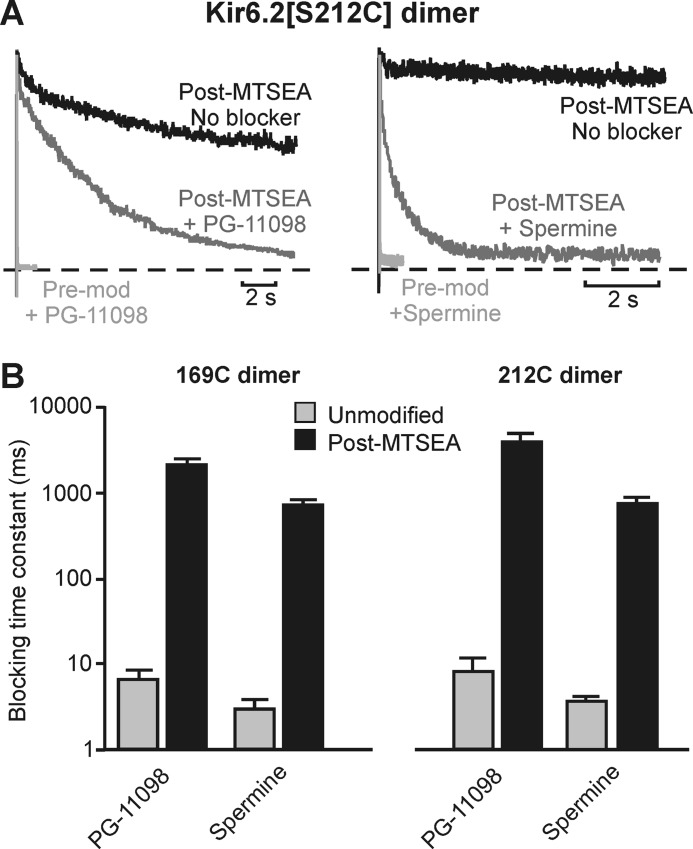
Steeply voltage-dependent inward rectification of Kir (inwardly rectifying potassium) channels arises from blockade by cytoplasmic polyamines. These polycationic blockers traverse a long (>70 Å) pore, displacing multiple permeant ions, en route to a high affinity binding site that remains loosely defined. We have scanned the effects of cysteine modification at multiple pore-lining positions on the blocking properties of a library of polyamine analogs, demonstrating that the effects of cysteine modification are position- and blocker-dependent. Specifically, introduction of positively charged adducts results in two distinct phenotypes: either disruption of blocker binding or generation of a barrier to blocker migration, in a consistent pattern that depends on both the length of the polyamine blocker and the position of the modified cysteine. These findings reveal important details about the chemical basis and specific location of high affinity polyamine binding.
Figures
References
-
- Lu Z. (2004) Mechanism of rectification in inward-rectifier K+ channels. Annu. Rev. Physiol. 66, 103–129 - PubMed
-
- Nichols C. G., Lopatin A. N. (1997) Inward rectifier potassium channels. Annu. Rev. Physiol. 59, 171–191 - PubMed
-
- Kuo A., Gulbis J. M., Antcliff J. F., Rahman T., Lowe E. D., Zimmer J., Cuthbertson J., Ashcroft F. M., Ezaki T., Doyle D. A. (2003) Crystal structure of the potassium channel KirBac1.1 in the closed state. Science 300, 1922–1926 - PubMed
-
- Lopatin A. N., Makhina E. N., Nichols C. G. (1994) Potassium channel block by cytoplasmic polyamines as the mechanism of intrinsic rectification. Nature 372, 366–369 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
