

Lineage tracing of Pf4-Cre marks hematopoietic stem cells and their progeny
- PMID: 23300543
- PMCID: PMC3531453
- DOI: 10.1371/journal.pone.0051361
Lineage tracing of Pf4-Cre marks hematopoietic stem cells and their progeny
Erratum in
- PLoS One. 2013;8(5). doi:10.1371/annotation/4c610387-c7f0-4330-b772-30ed52a07547. Guitart, Amelie [corrected to Guitart, Amelie V]
Abstract
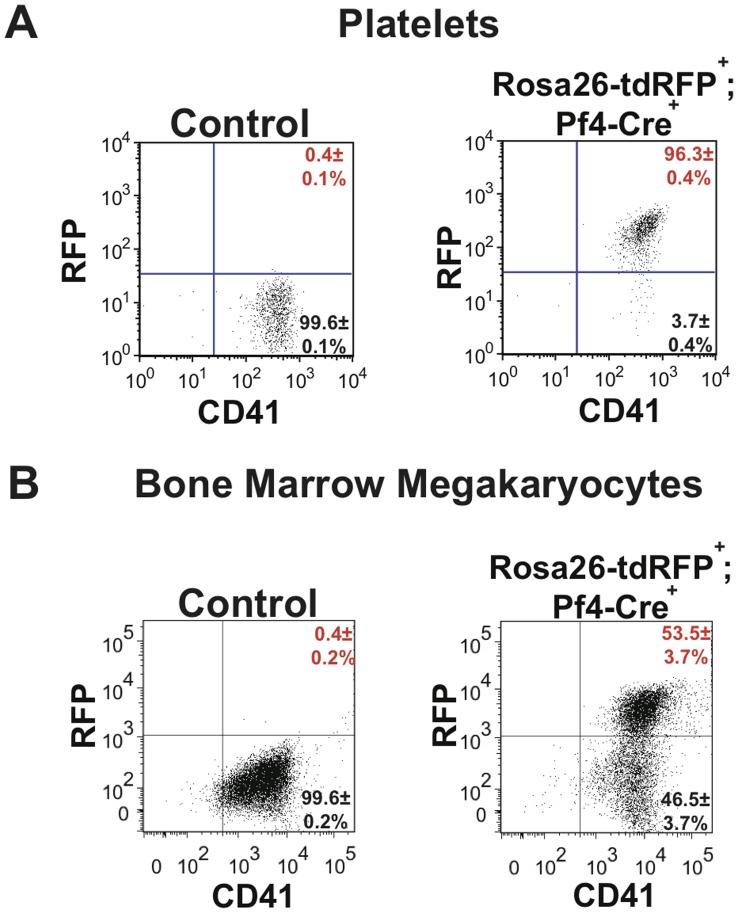
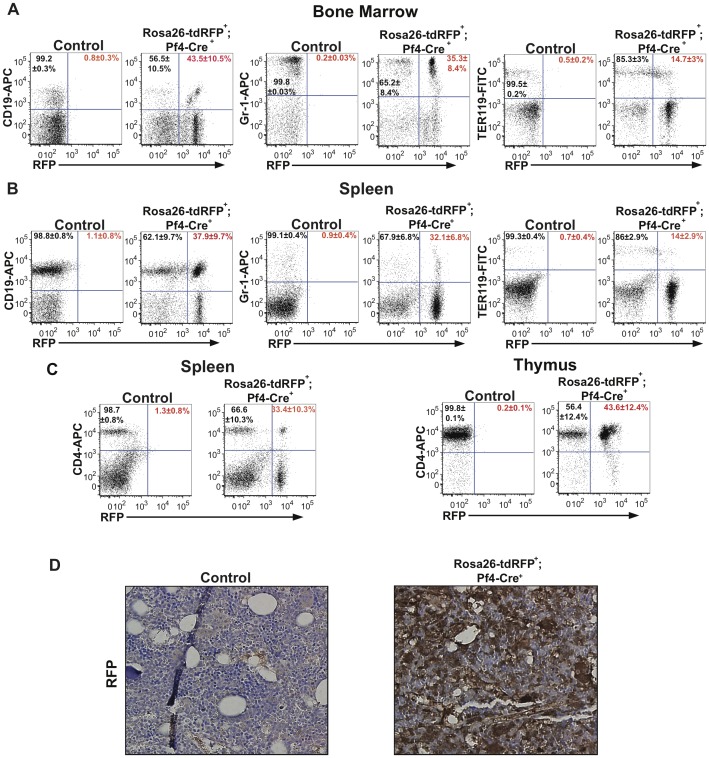
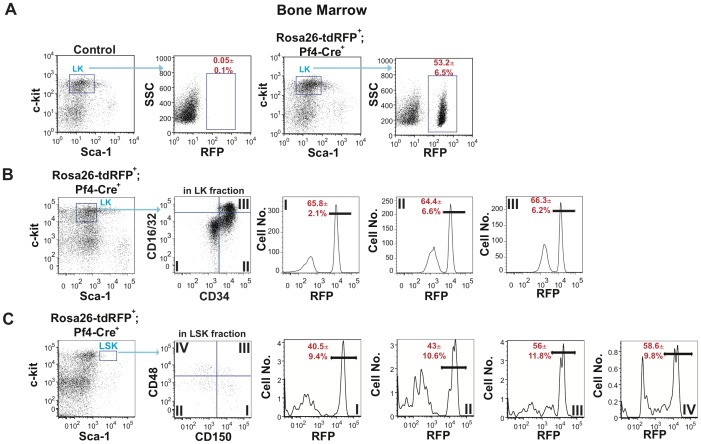
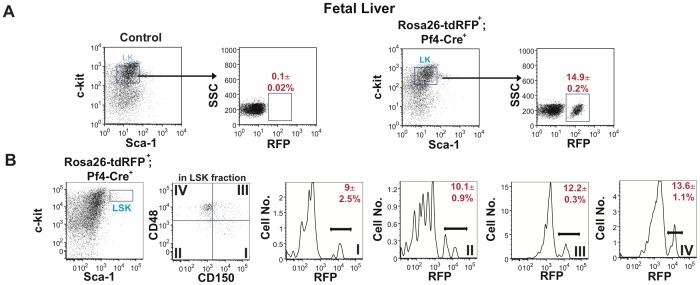
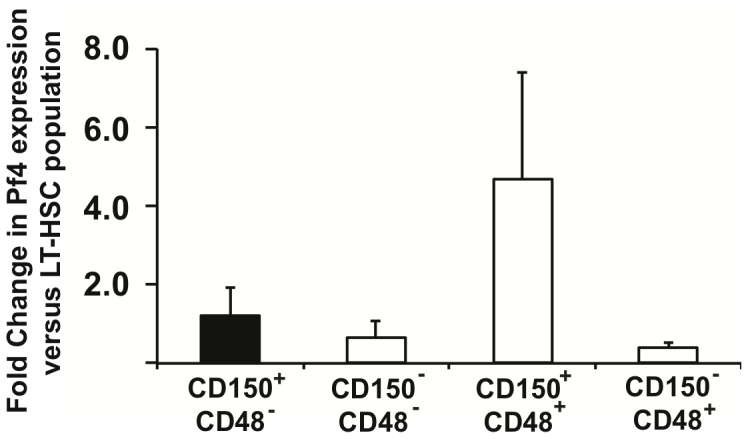
The development of a megakaryocyte lineage specific Cre deleter, using the Pf4 (CXCL4) promoter (Pf4-Cre), was a significant step forward in the specific analysis of platelet and megakaryocyte cell biology. However, in the present study we have employed a sensitive reporter-based approach to demonstrate that Pf4-Cre also recombines in a significant proportion of both fetal liver and bone marrow hematopoietic stem cells (HSCs), including the most primitive fraction containing the long-term repopulating HSCs. Consequently, we demonstrate that Pf4-Cre activity is not megakaryocyte lineage-specific but extends to other myeloid and lymphoid lineages at significant levels between 15-60%. Finally, we show for the first time that Pf4 transcripts are present in adult HSCs and primitive hematopoietic progenitor cells. These results have fundamental implications for the use of the Pf4-Cre mouse model and for our understanding of a possible role for Pf4 in the development of the hematopoietic lineage.
Conflict of interest statement
Figures
References
-
- Bockamp E, Sprengel R, Eshkind L, Lehmann T, Braun JM, et al. (2008) Conditional transgenic mouse models: from the basics to genome-wide sets of knockouts and current studies of tissue regeneration. Regenerative medicine 3: 217–235. - PubMed
-
- de Boer J, Williams A, Skavdis G, Harker N, Coles M, et al. (2003) Transgenic mice with hematopoietic and lymphoid specific expression of Cre. European journal of immunology 33: 314–325. - PubMed
-
- Hayashi S, McMahon AP (2002) Efficient recombination in diverse tissues by a tamoxifen-inducible form of Cre: a tool for temporally regulated gene activation/inactivation in the mouse. Developmental biology 244: 305–318. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
