

Transcriptional program of ciliated epithelial cells reveals new cilium and centrosome components and links to human disease
- PMID: 23300604
- PMCID: PMC3534086
- DOI: 10.1371/journal.pone.0052166
Transcriptional program of ciliated epithelial cells reveals new cilium and centrosome components and links to human disease
Abstract
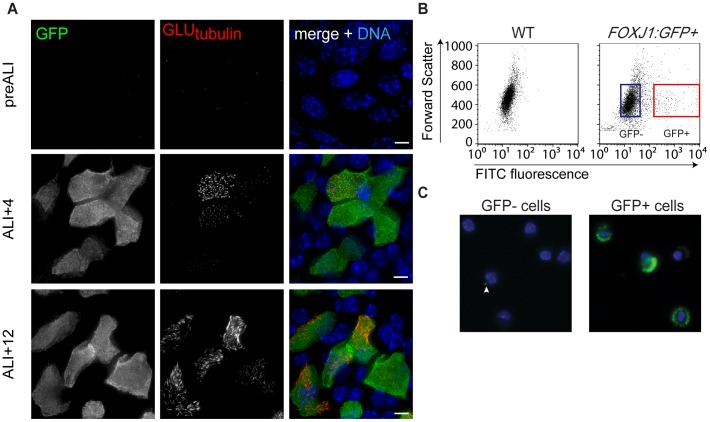
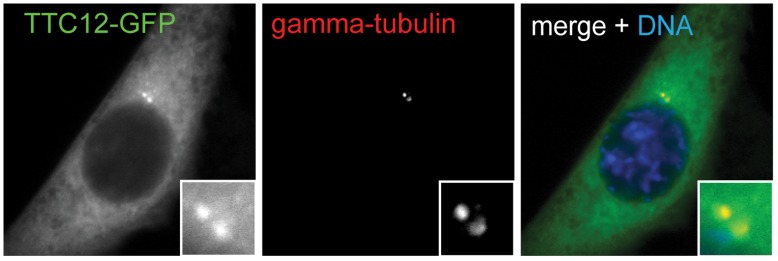
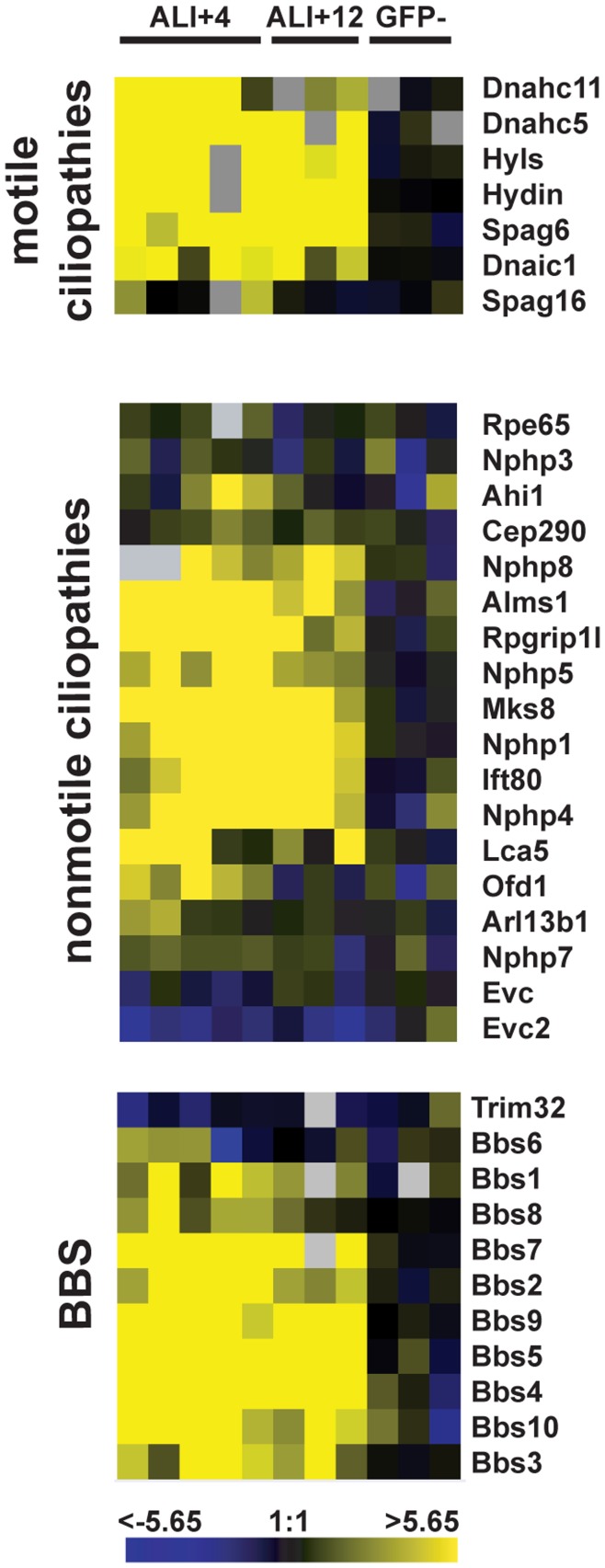
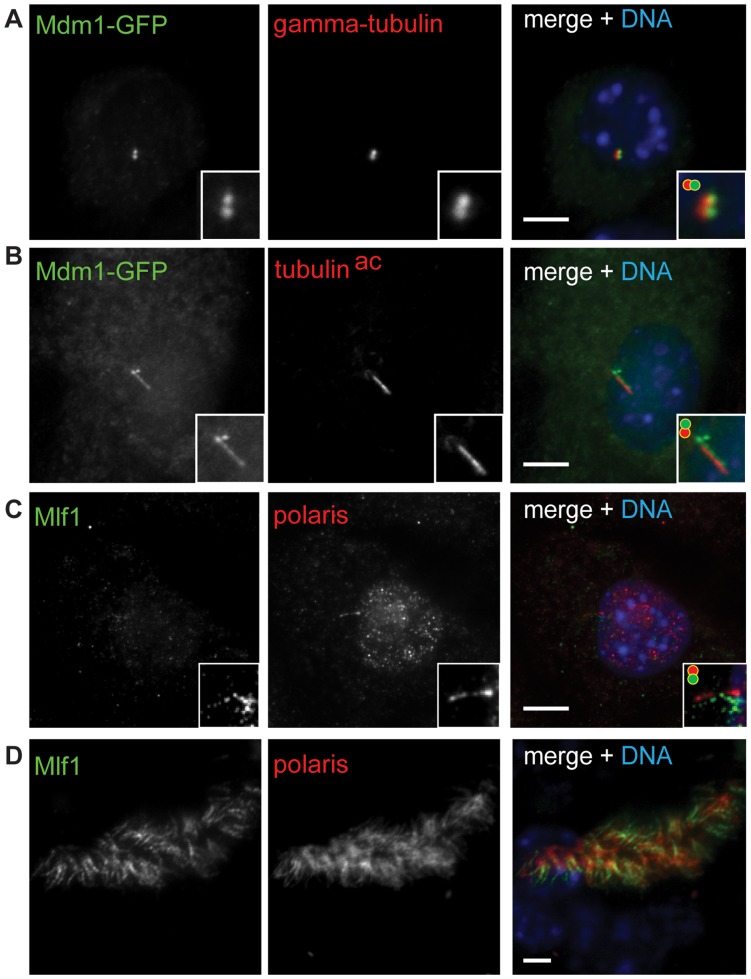
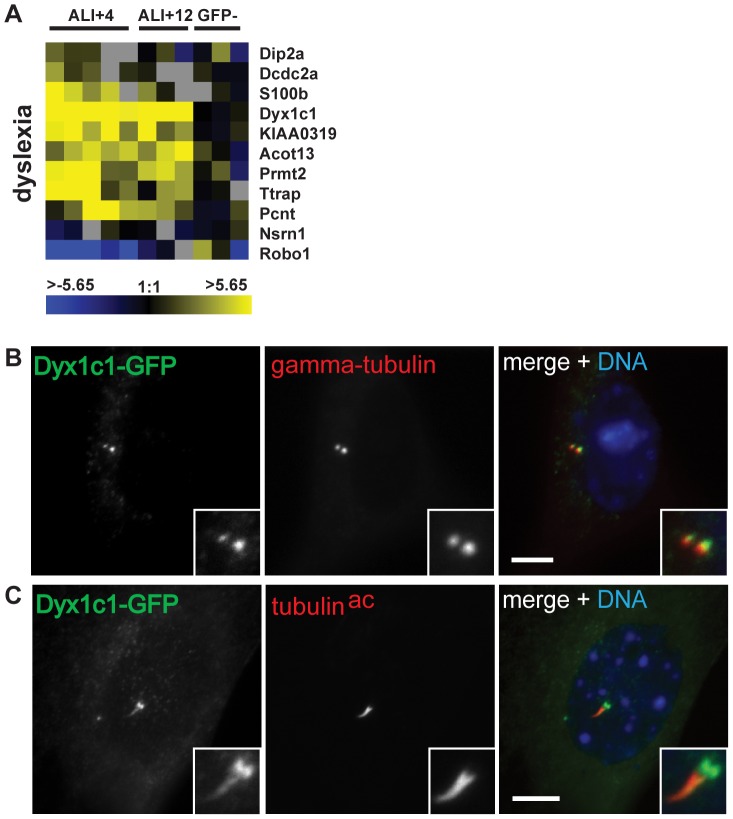
Defects in the centrosome and cilium are associated with a set of human diseases having diverse phenotypes. To further characterize the components that define the function of these organelles we determined the transcriptional profile of multiciliated tracheal epithelial cells. Cultures of mouse tracheal epithelial cells undergoing differentiation in vitro were derived from mice expressing GFP from the ciliated-cell specific FOXJ1 promoter (FOXJ1:GFP). The transcriptional profile of ciliating GFP+ cells from these cultures was defined at an early and a late time point during differentiation and was refined by subtraction of the profile of the non-ciliated GFP- cells. We identified 649 genes upregulated early, when most cells were forming basal bodies, and 73 genes genes upregulated late, when most cells were fully ciliated. Most, but not all, of known centrosome proteins are transcriptionally upregulated early, particularly Plk4, a master regulator of centriole formation. We found that three genes associated with human disease states, Mdm1, Mlf1, and Dyx1c1, are upregulated during ciliogenesis and localize to centrioles and cilia. This transcriptome for mammalian multiciliated epithelial cells identifies new candidate centrosome and cilia proteins, highlights similarities between components of motile and primary cilia, and identifies new links between cilia proteins and human disease.
Conflict of interest statement
Figures



References
-
- Vincensini L, Blisnick T, Bastin P (2011) 1001 model organisms to study cilia and flagella. Biol Cell 103: 109–130. - PubMed
-
- Fisch C, Dupuis-Williams P (2011) Ultrastructure of cilia and flagella - back to the future! Biol Cell. 103: 249–270. - PubMed
-
- Christensen ST, Guerra CF, Awan A, Wheatley DN, Satir P (2003) Insulin receptor-like proteins in Tetrahymena thermophila ciliary membranes. Curr Biol 13: R50–52. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
