

Proteomic analysis of lipid droplets from Caco-2/TC7 enterocytes identifies novel modulators of lipid secretion
- PMID: 23301014
- PMCID: PMC3534623
- DOI: 10.1371/journal.pone.0053017
Proteomic analysis of lipid droplets from Caco-2/TC7 enterocytes identifies novel modulators of lipid secretion
Abstract
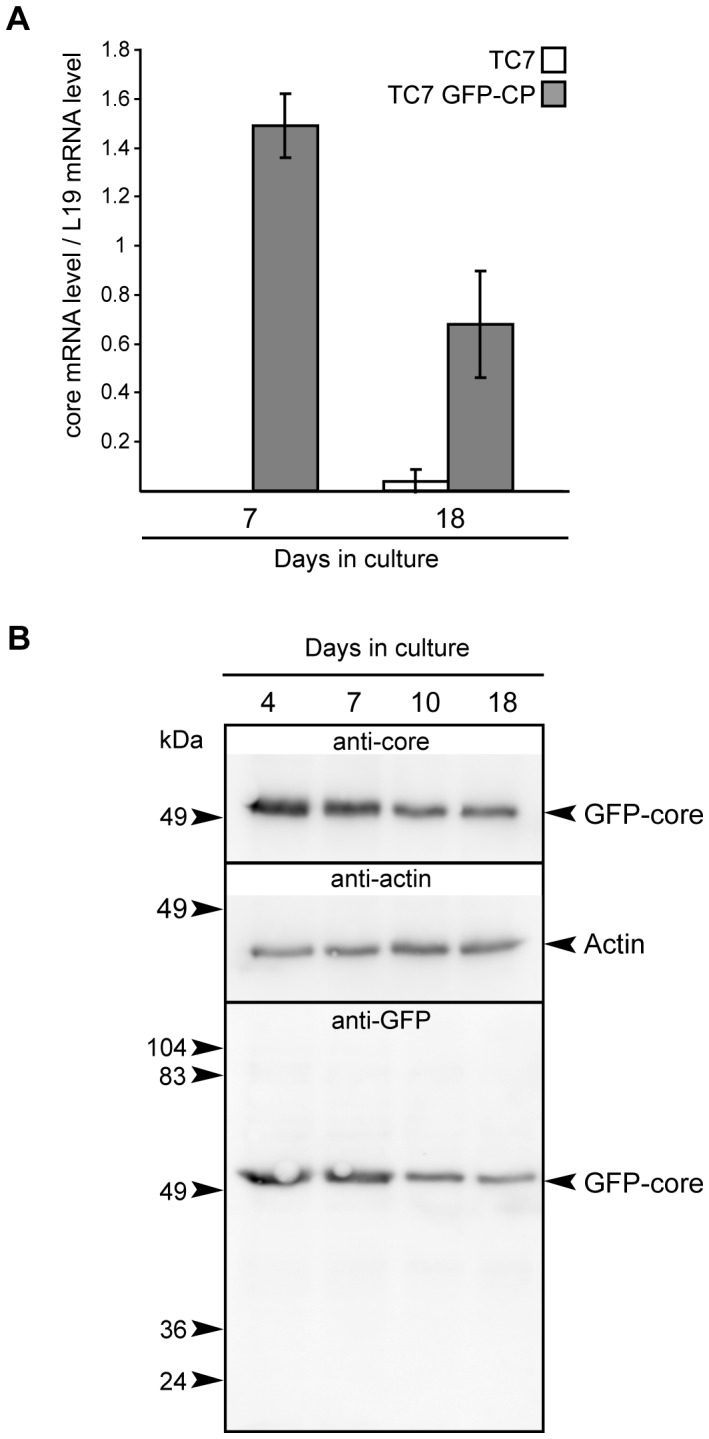
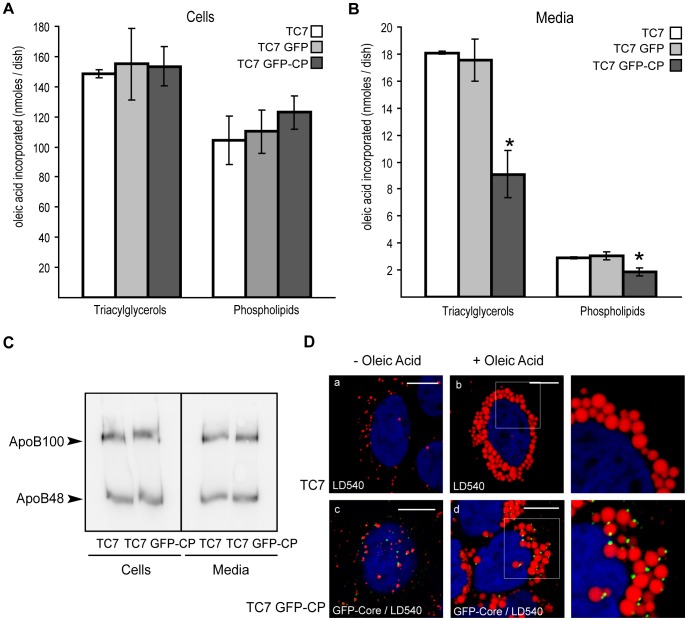
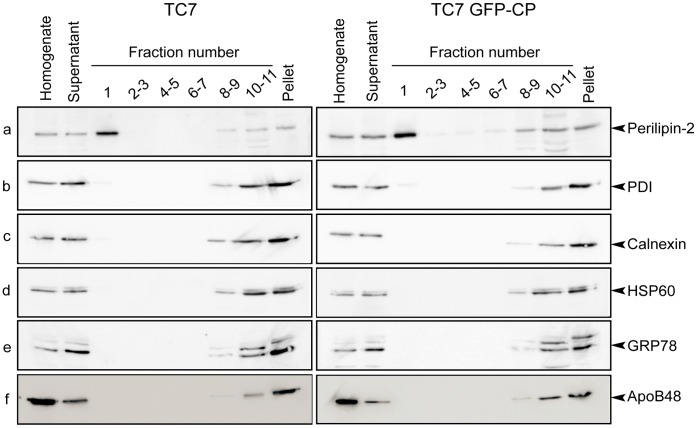
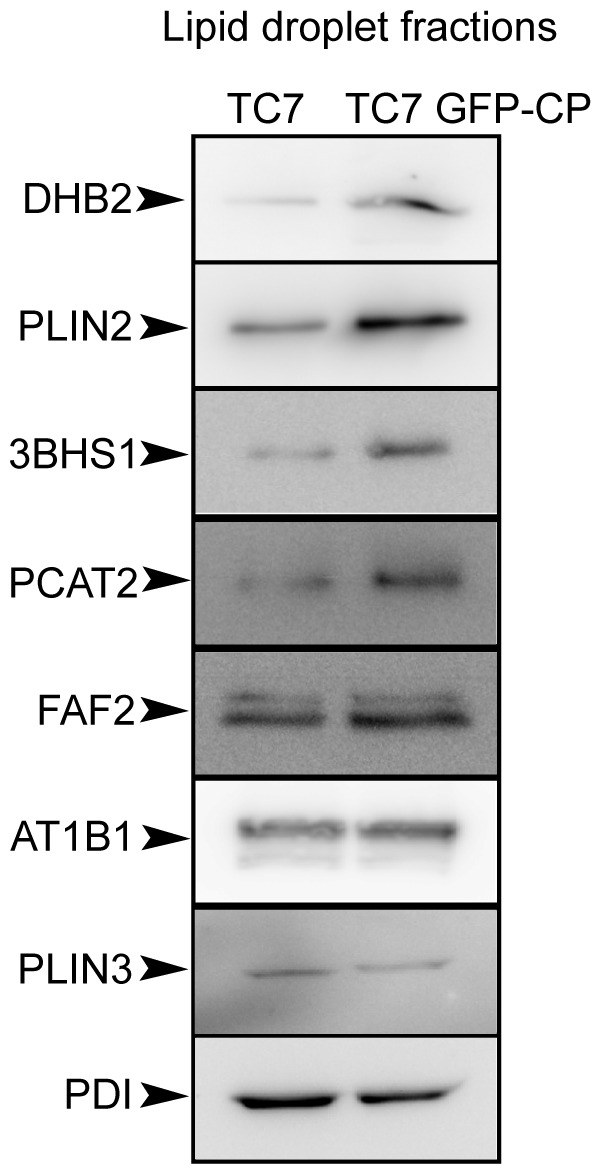
In enterocytes, the dynamic accumulation and depletion of triacylglycerol (TAG) in lipid droplets (LD) during fat absorption suggests that cytosolic LD-associated TAG contribute to TAG-rich lipoprotein (TRL) production. To get insight into the mechanisms controlling the storage/secretion balance of TAG, we used as a tool hepatitis C virus core protein, which localizes onto LDs, and thus may modify their protein coat and decrease TRL secretion. We compared the proteome of LD fractions isolated from Caco-2/TC7 enterocytes expressing or not hepatitis C virus core protein by a differential proteomic approach (isobaric tag for relative and absolute quantitation (iTRAQ) labeling coupled with liquid chromatography and tandem mass spectrometry). We identified 42 proteins, 21 being involved in lipid metabolism. Perilipin-2/ADRP, which is suggested to stabilize long term-stored TAG, was enriched in LD fractions isolated from Caco-2/TC7 expressing core protein while perilipin-3/TIP47, which is involved in LD synthesis from newly synthesized TAG, was decreased. Endoplasmic reticulum-associated proteins were strongly decreased, suggesting reduced interactions between LD and endoplasmic reticulum, where TRL assembly occurs. For the first time, we show that 17β-hydroxysteroid dehydrogenase 2 (DHB2), which catalyzes the conversion of 17-keto to 17 β-hydroxysteroids and which was the most highly enriched protein in core expressing cells, is localized to LD and interferes with TAG secretion, probably through its capacity to inactivate testosterone. Overall, we identified potential new players of lipid droplet dynamics, which may be involved in the balance between lipid storage and secretion, and may be altered in enterocytes in pathological conditions such as insulin resistance, type II diabetes and obesity.
Conflict of interest statement
Figures
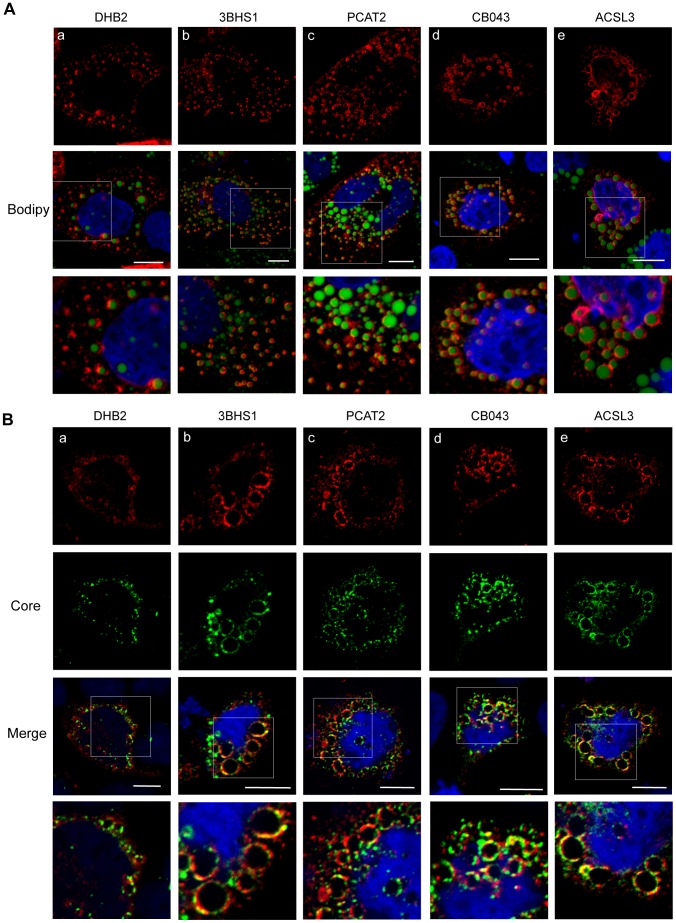
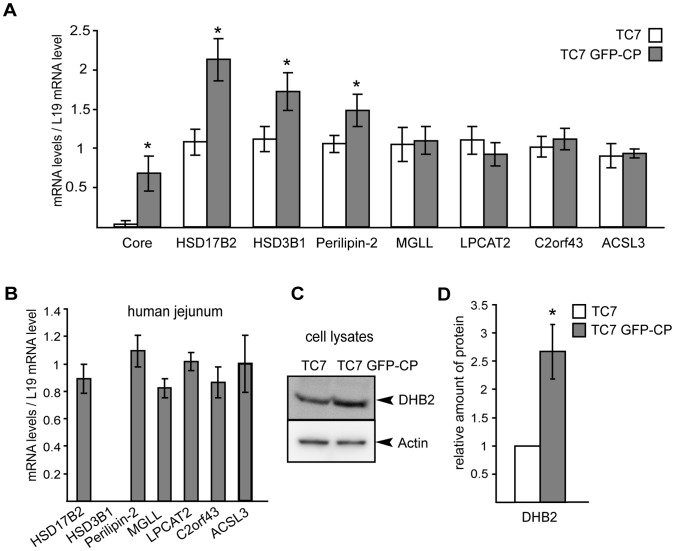
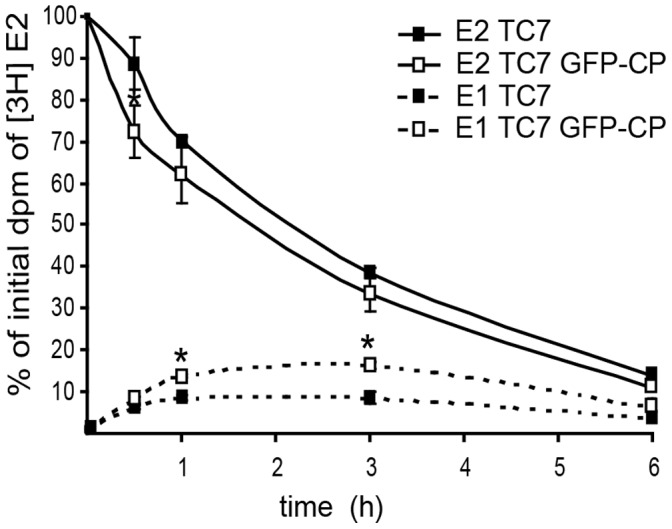
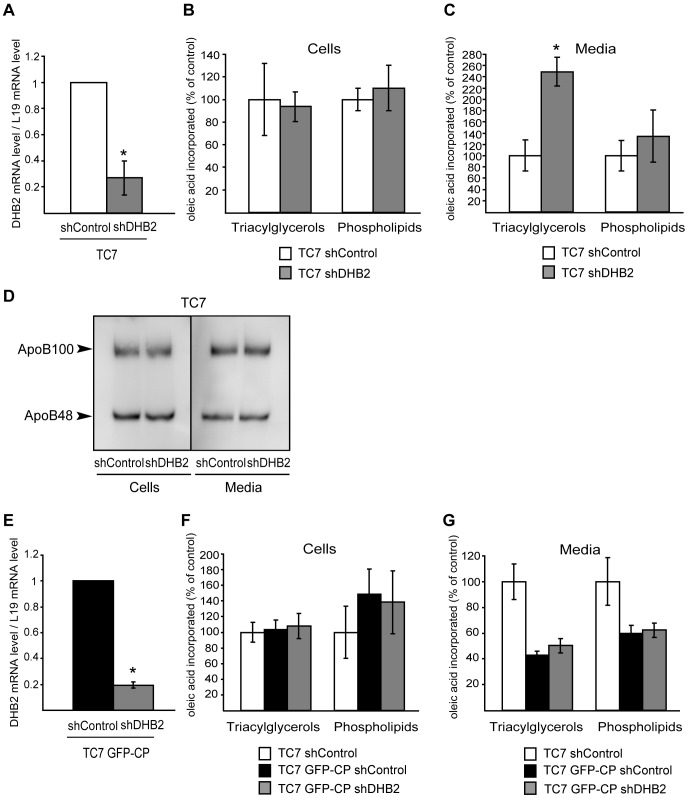
References
-
- Murphy DJ, Vance J (1999) Mechanisms of lipid-body formation. Trends Biochem Sci 24: 109–115. - PubMed
-
- Ohsaki Y, Cheng J, Suzuki M, Shinohara Y, Fujita A, et al. (2009) Biogenesis of cytoplasmic lipid droplets: From the lipid ester globule in the membrane to the visible structure. Biochim Biophys Acta 1791: 399–407. - PubMed
-
- Lehner R, Lian J, Quiroga AD (2012) Lumenal lipid metabolism: implications for lipoprotein assembly. Arterioscler Thromb Vasc Biol 32: 1087–1093. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
