

RNA-Seq profiling of spinal cord motor neurons from a presymptomatic SOD1 ALS mouse
- PMID: 23301088
- PMCID: PMC3536741
- DOI: 10.1371/journal.pone.0053575
RNA-Seq profiling of spinal cord motor neurons from a presymptomatic SOD1 ALS mouse
Abstract
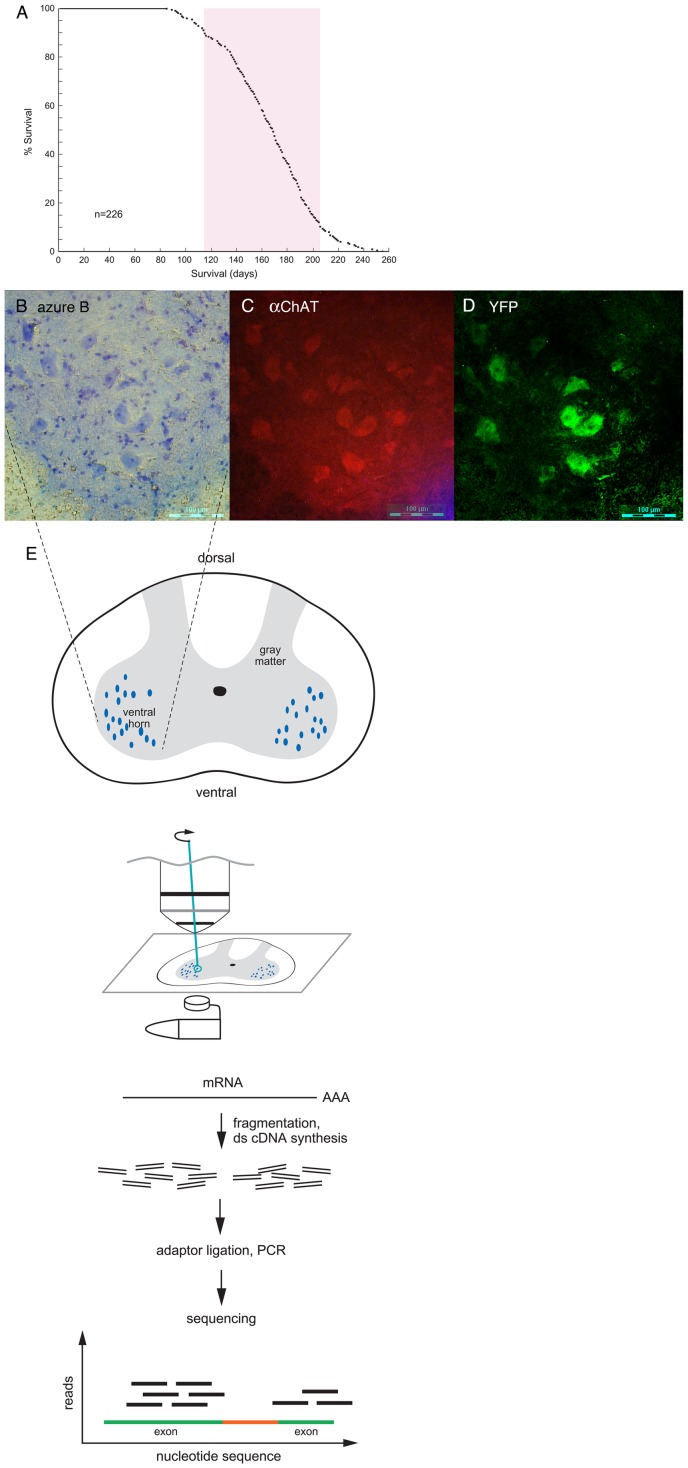
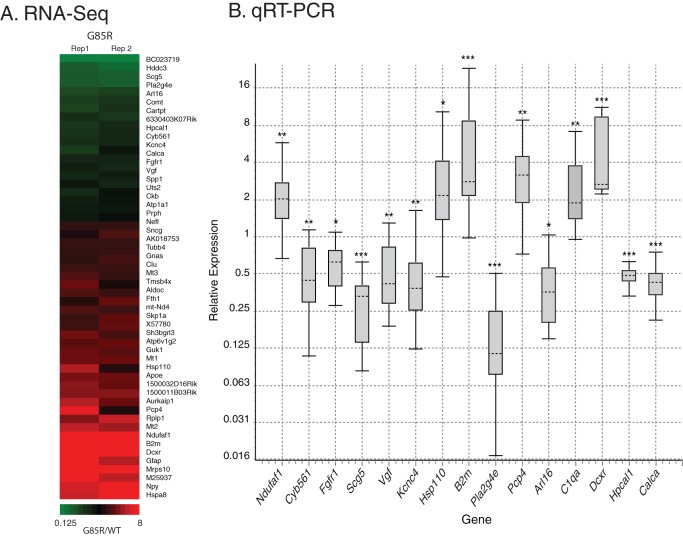
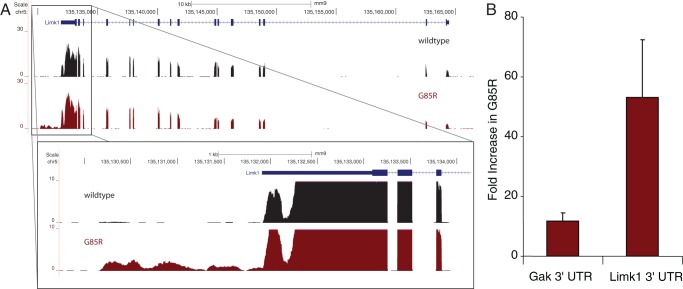
Mechanisms involved with degeneration of motor neurons in amyotrophic lateral sclerosis (ALS; Lou Gehrig's Disease) are poorly understood, but genetically inherited forms, comprising ~10% of the cases, are potentially informative. Recent observations that several inherited forms of ALS involve the RNA binding proteins TDP43 and FUS raise the question as to whether RNA metabolism is generally disturbed in ALS. Here we conduct whole transcriptome profiling of motor neurons from a mouse strain, transgenic for a mutant human SOD1 (G85R SOD1-YFP), that develops symptoms of ALS and paralyzes at 5-6 months of age. Motor neuron cell bodies were laser microdissected from spinal cords at 3 months of age, a time when animals were presymptomatic but showed aggregation of the mutant protein in many lower motor neuron cell bodies and manifested extensive neuromuscular junction morphologic disturbance in their lower extremities. We observed only a small number of transcripts with altered expression levels or splicing in the G85R transgenic compared to age-matched animals of a wild-type SOD1 transgenic strain. Our results indicate that a major disturbance of polyadenylated RNA metabolism does not occur in motor neurons of mutant SOD1 mice, suggesting that the toxicity of the mutant protein lies at the level of translational or post-translational effects.
Conflict of interest statement
Figures
References
-
- Rosen DR, Siddique T, Patterson D, Figlewicz DA, Sapp P, et al. (1993) Mutations in Cu/Zn superoxide dismutase gene are associated with familial amyotrophic lateral sclerosis. Nature 362: 59–62. - PubMed
-
- Hart PJ (2006) Pathogenic superoxide dismutase structure, folding, aggregation and turnover. Curr Op Chem Biol 10: 131–138. - PubMed
-
- Maruyama H, Morino H, Ito H, Izumi Y, Kato H, et al. (2010) Mutations of optineurin in amyotrophic lateral sclerosis. Nature 465: 223–226. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
