

Tetraspanin CD151 mediates papillomavirus type 16 endocytosis
- PMID: 23302890
- PMCID: PMC3592167
- DOI: 10.1128/JVI.02906-12
Tetraspanin CD151 mediates papillomavirus type 16 endocytosis
Abstract
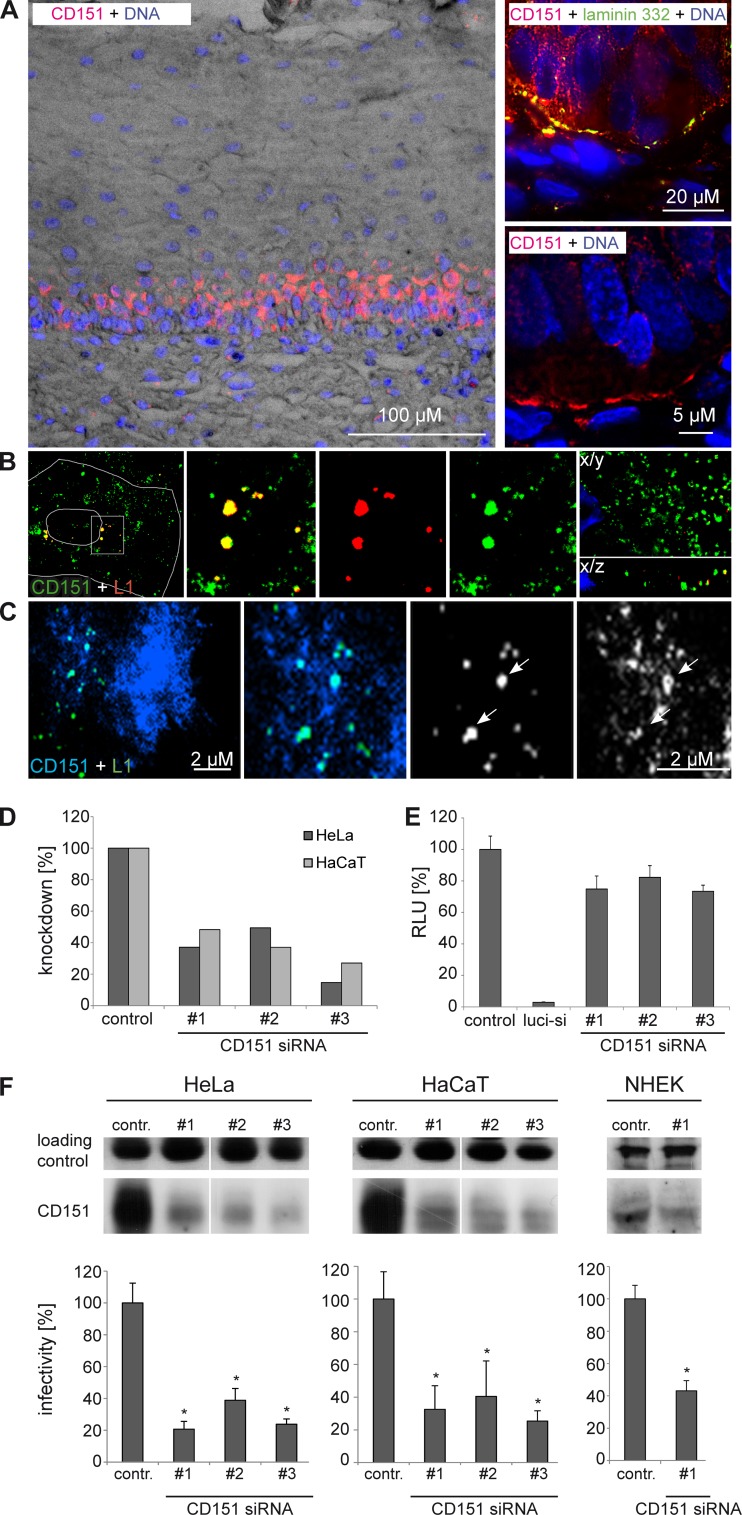
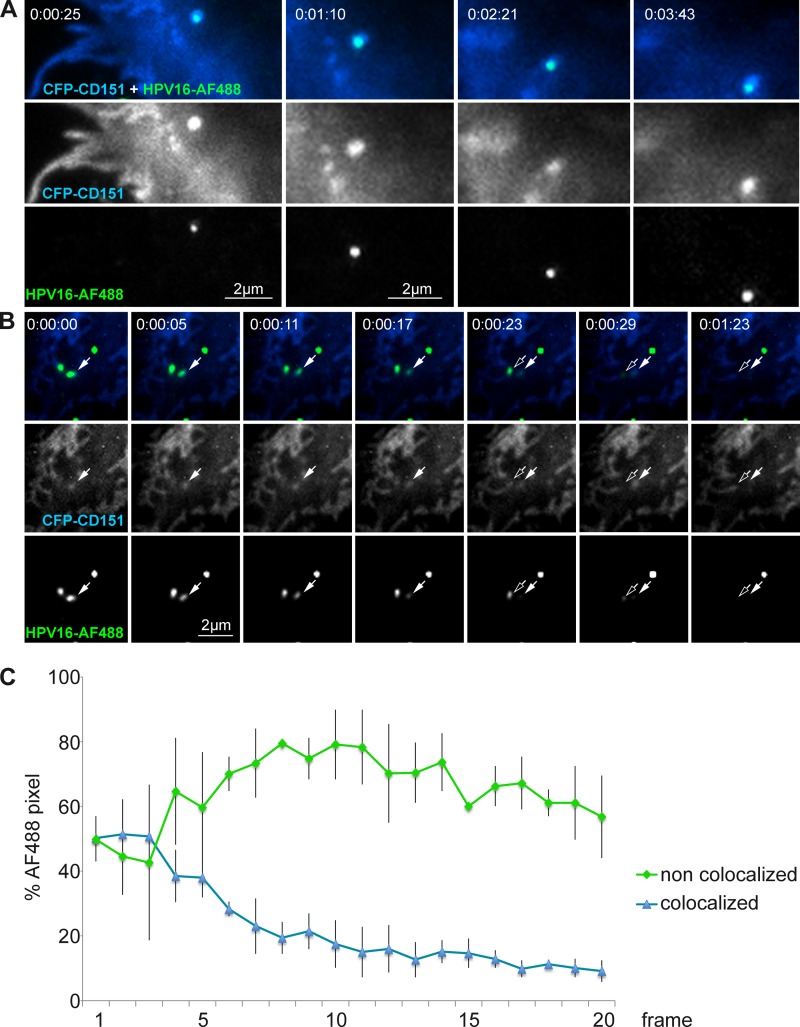
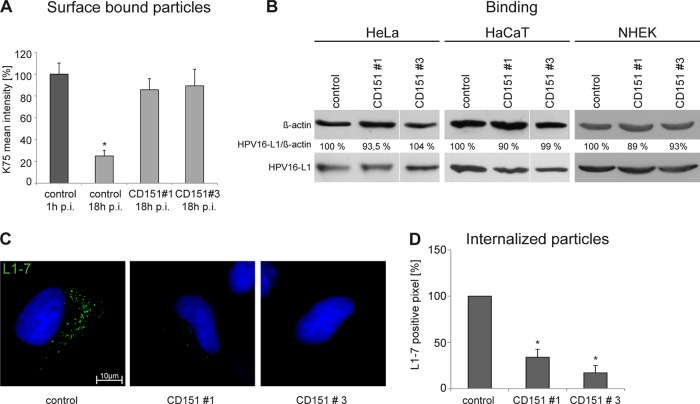
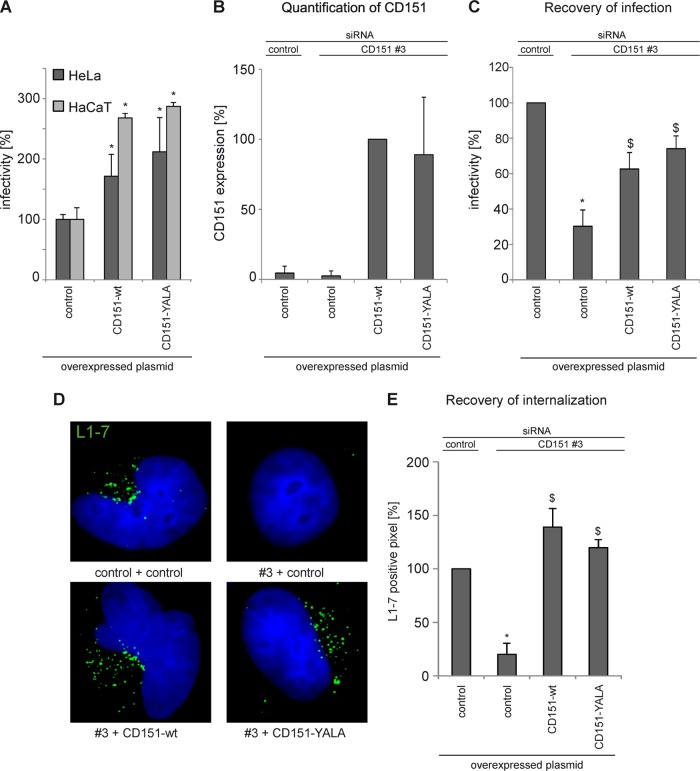
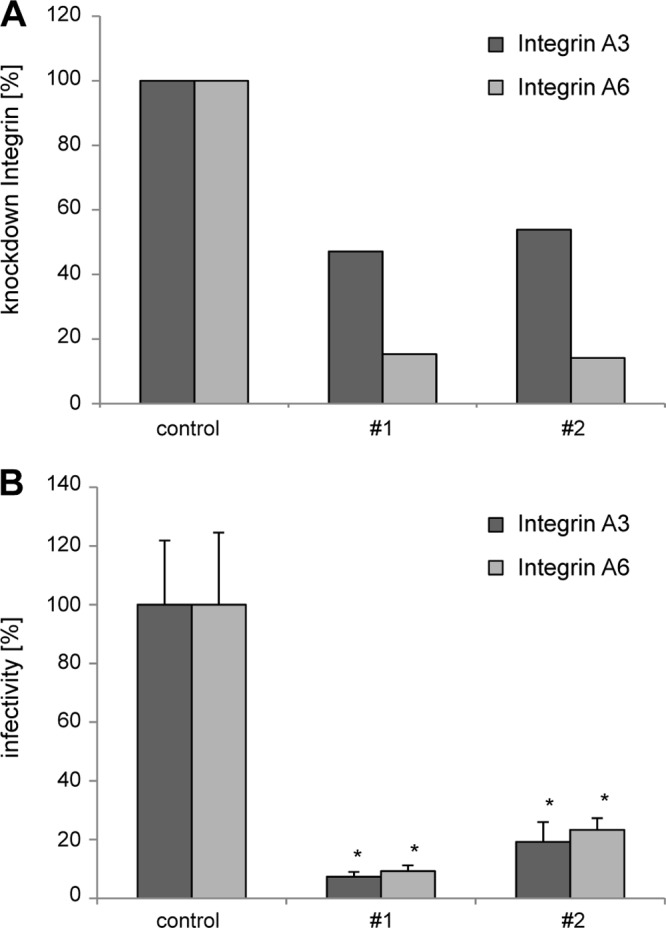
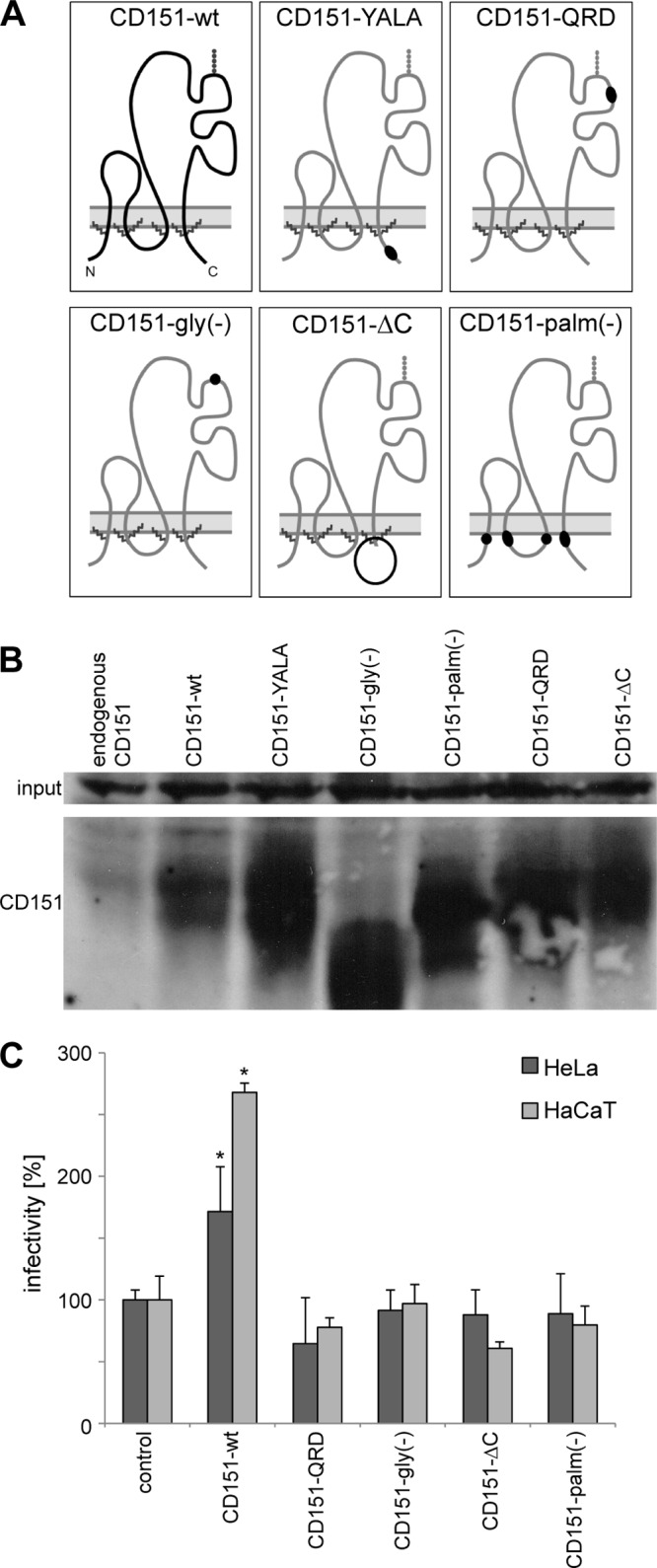
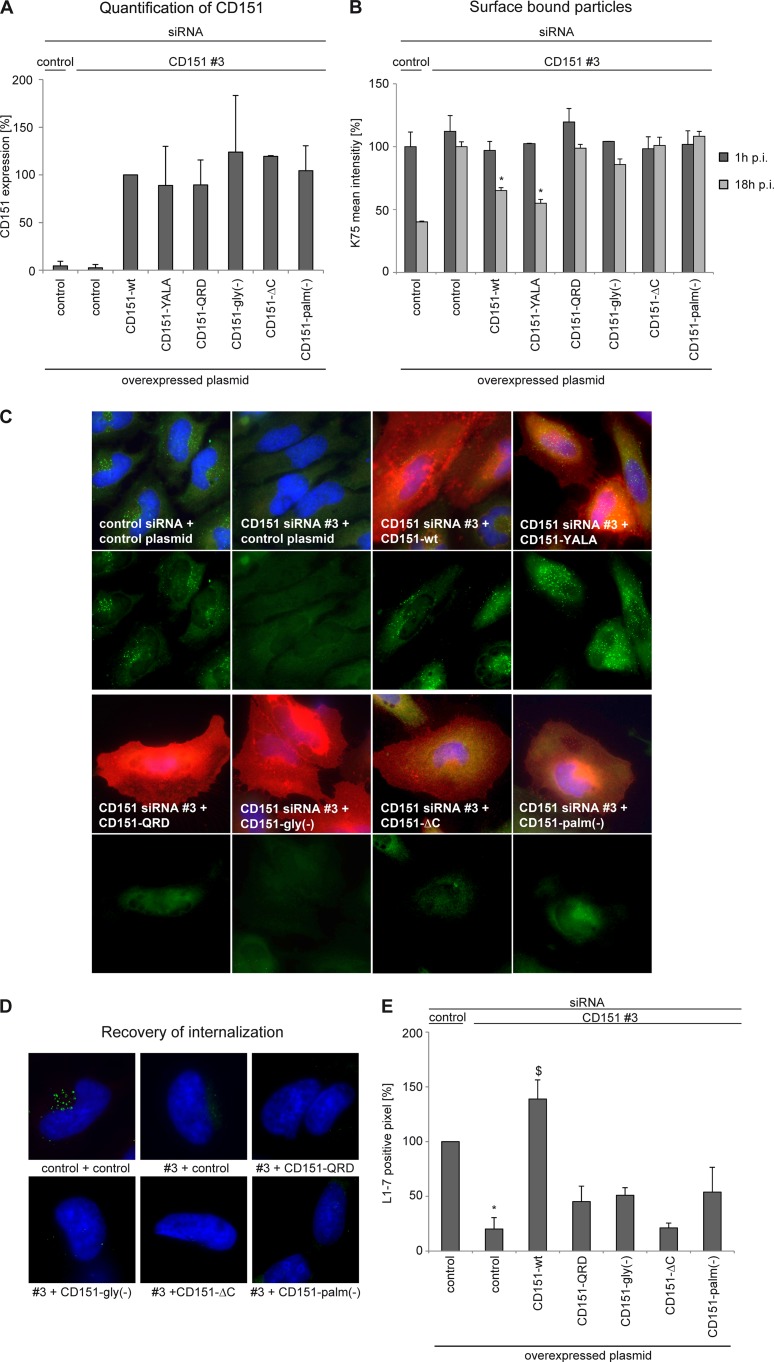
Human papillomavirus type 16 (HPV16) is the primary etiologic agent for cervical cancer. The infectious entry of HPV16 into cells occurs via a so-far poorly characterized clathrin- and caveolin-independent endocytic pathway, which involves tetraspanin proteins and actin. In this study, we investigated the specific role of the tetraspanin CD151 in the early steps of HPV16 infection. We show that surface-bound HPV16 moves together with CD151 within the plane of the membrane before they cointernalize into endosomes. Depletion of endogenous CD151 did not affect binding of viral particles to cells but resulted in reduction of HPV16 endocytosis. HPV16 uptake is dependent on the C-terminal cytoplasmic region of CD151 but does not require its tyrosine-based sorting motif. Reexpression of the wild-type CD151 but not mutants affecting integrin functions restored virus internalization in CD151-depleted cells. Accordingly, short interfering RNA (siRNA) gene knockdown experiments confirmed that CD151-associated integrins (i.e., α3β1 and α6β1/4) are involved in HPV16 infection. Furthermore, palmitoylation-deficient CD151 did not support HPV16 cell entry. These data show that complex formation of CD151 with laminin-binding integrins and integration of the complex into tetraspanin-enriched microdomains are critical for HPV16 endocytosis.
Figures
References
-
- Chen X, Garcea R, Goldberg I, Casini G, Harrison S. 2000. Structure of small virus-like particles assembled from the L1 protein of human papillomavirus 16. Mol. Cell 5:557–567 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
