

Alarming features: birds use specific acoustic properties to identify heterospecific alarm calls
- PMID: 23303539
- PMCID: PMC3574322
- DOI: 10.1098/rspb.2012.2539
Alarming features: birds use specific acoustic properties to identify heterospecific alarm calls
Abstract
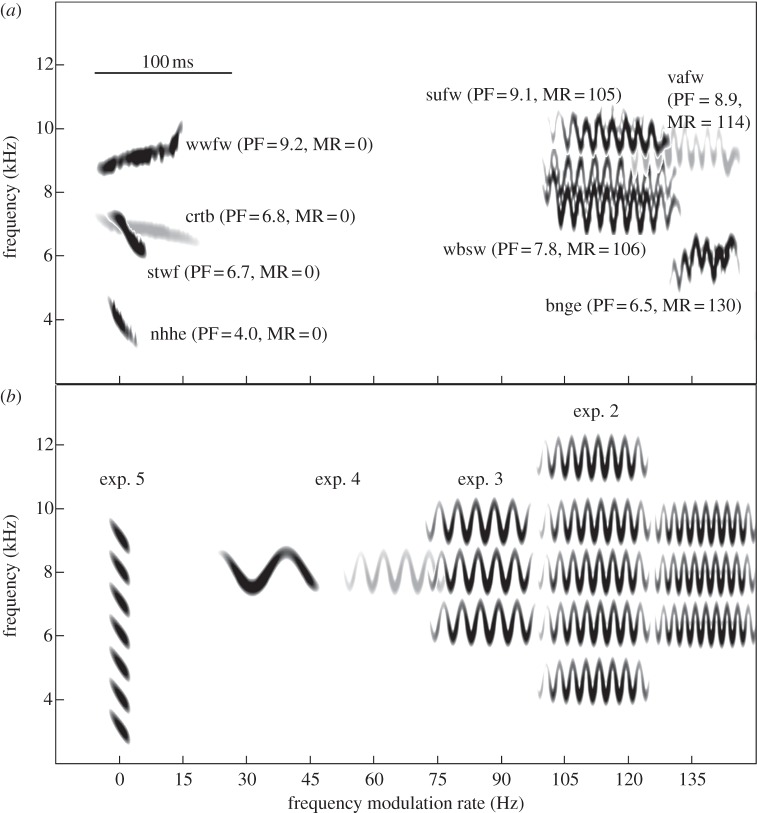
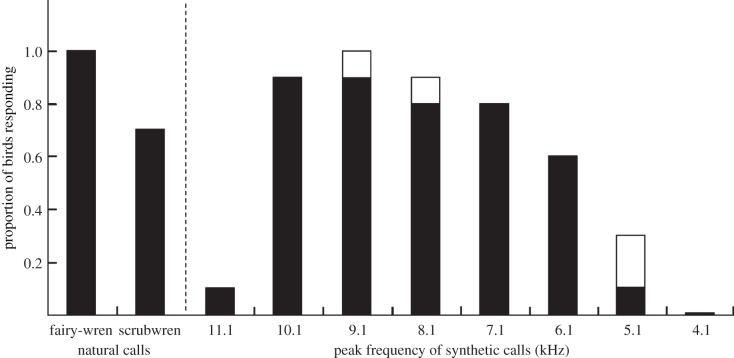
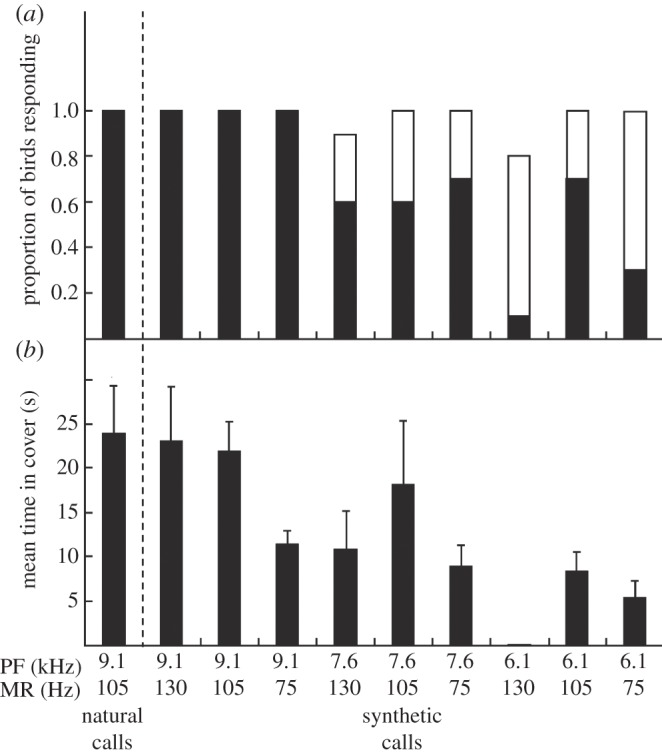
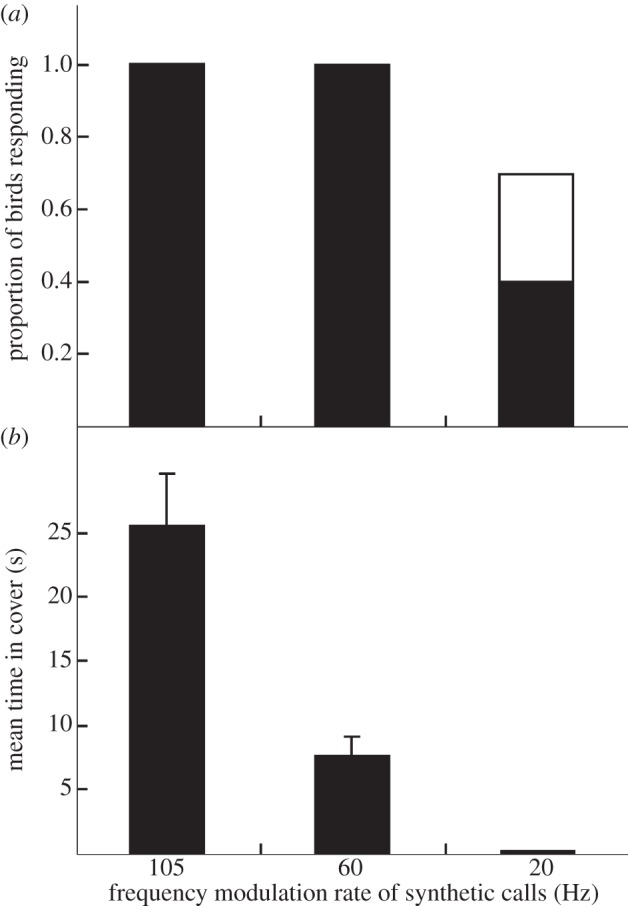
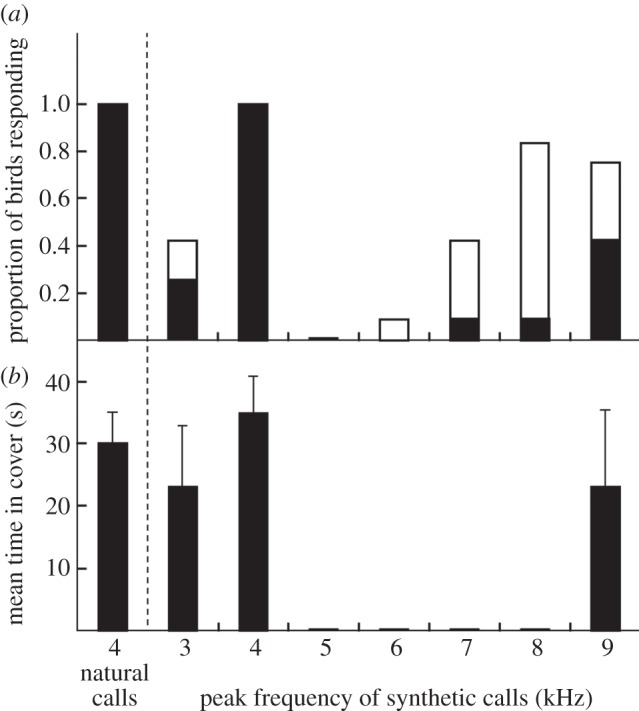
Vertebrates that eavesdrop on heterospecific alarm calls must distinguish alarms from sounds that can safely be ignored, but the mechanisms for identifying heterospecific alarm calls are poorly understood. While vertebrates learn to identify heterospecific alarms through experience, some can also respond to unfamiliar alarm calls that are acoustically similar to conspecific alarm calls. We used synthetic calls to test the role of specific acoustic properties in alarm call identification by superb fairy-wrens, Malurus cyaneus. Individuals fled more often in response to synthetic calls with peak frequencies closer to those of conspecific calls, even if other acoustic features were dissimilar to that of fairy-wren calls. Further, they then spent more time in cover following calls that had both peak frequencies and frequency modulation rates closer to natural fairy-wren means. Thus, fairy-wrens use similarity in specific acoustic properties to identify alarms and adjust a two-stage antipredator response. Our study reveals how birds respond to heterospecific alarm calls without experience, and, together with previous work using playback of natural calls, shows that both acoustic similarity and learning are important for interspecific eavesdropping. More generally, this study reconciles contrasting views on the importance of alarm signal structure and learning in recognition of heterospecific alarms.
Figures
Similar articles
-
Noise constrains heterospecific eavesdropping more than conspecific reception of alarm calls.Biol Lett. 2024 Jan;20(1):20230410. doi: 10.1098/rsbl.2023.0410. Epub 2024 Jan 17. Biol Lett. 2024. PMID: 38228188 Free PMC article.
-
Wild birds learn to eavesdrop on heterospecific alarm calls.Curr Biol. 2015 Aug 3;25(15):2047-50. doi: 10.1016/j.cub.2015.06.028. Epub 2015 Jul 16. Curr Biol. 2015. PMID: 26190077
-
Birds Learn Socially to Recognize Heterospecific Alarm Calls by Acoustic Association.Curr Biol. 2018 Aug 20;28(16):2632-2637.e4. doi: 10.1016/j.cub.2018.06.013. Epub 2018 Aug 2. Curr Biol. 2018. PMID: 30078559
-
Eavesdropping on heterospecific alarm calls: from mechanisms to consequences.Biol Rev Camb Philos Soc. 2015 May;90(2):560-86. doi: 10.1111/brv.12122. Epub 2014 Jun 11. Biol Rev Camb Philos Soc. 2015. PMID: 24917385 Review.
-
Pharmacological and behavioral characteristics of 22 kHz alarm calls in rats.Neurosci Biobehav Rev. 2001 Dec;25(7-8):611-7. doi: 10.1016/s0149-7634(01)00058-6. Neurosci Biobehav Rev. 2001. PMID: 11801286 Review.
Cited by
-
Multiple adaptive and non-adaptive processes determine responsiveness to heterospecific alarm calls in African savannah herbivores.Proc Biol Sci. 2018 Jul 4;285(1882):20172676. doi: 10.1098/rspb.2017.2676. Proc Biol Sci. 2018. PMID: 30051827 Free PMC article.
-
Noise constrains heterospecific eavesdropping more than conspecific reception of alarm calls.Biol Lett. 2024 Jan;20(1):20230410. doi: 10.1098/rsbl.2023.0410. Epub 2024 Jan 17. Biol Lett. 2024. PMID: 38228188 Free PMC article.
-
Olfactory learning and memory in the greater short-nosed fruit bat Cynopterus sphinx: the influence of conspecifics distress calls.J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2021 Sep;207(5):667-679. doi: 10.1007/s00359-021-01505-2. Epub 2021 Aug 23. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2021. PMID: 34426872
-
Learning and signal copying facilitate communication among bird species.Proc Biol Sci. 2013 Feb 27;280(1757):20123070. doi: 10.1098/rspb.2012.3070. Print 2013 Apr 22. Proc Biol Sci. 2013. PMID: 23446529 Free PMC article.
-
Social learning and culture in birds: emerging patterns and relevance to conservation.Philos Trans R Soc Lond B Biol Sci. 2025 May;380(1925):20240128. doi: 10.1098/rstb.2024.0128. Epub 2025 May 1. Philos Trans R Soc Lond B Biol Sci. 2025. PMID: 40308131 Free PMC article. Review.
References
-
- Vitousek MN, Adelman JS, Gregory NC, St Clair JJH. 2007. Heterospecific alarm call recognition in a non-vocal reptile. Biol. Lett. 3, 632–63410.1098/rsbl.2007.0443 (doi:10.1098/rsbl.2007.0443) - DOI - DOI - PMC - PubMed
-
- Ito R, Mori A. 2010. Vigilance against predators induced by eavesdropping on heterospecific alarm calls in a non-vocal lizard Oplurus cuvieri cuvieri (Reptilia: Iguania). Proc. R. Soc. B 277, 1275–128010.1098/rspb.2009.2047 (doi:10.1098/rspb.2009.2047) - DOI - DOI - PMC - PubMed
-
- Nuechterlein GL. 1981. ‘Information parasitism’ in mixed colonies of western grebes and Forster's terns. Anim. Behav. 29, 985–98910.1016/S0003-3472(81)80051-6 (doi:10.1016/S0003-3472(81)80051-6) - DOI - DOI
-
- Magrath RD, Pitcher BJ, Gardner JL. 2007. A mutual understanding? Interspecific responses by birds to each other's aerial alarm calls. Behav. Ecol. 18, 944–95110.1093/beheco/arm063 (doi:10.1093/beheco/arm063) - DOI - DOI
-
- Forsman JT, Monkkonen M. 2001. Responses by breeding birds to heterospecific song and mobbing call playbacks under varying predation risk. Anim. Behav. 62, 1067–107310.1006/anbe.2001.1856 (doi:10.1006/anbe.2001.1856) - DOI - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
