

Oxygen-dependent expression of cytochrome c oxidase subunit 4-2 gene expression is mediated by transcription factors RBPJ, CXXC5 and CHCHD2
- PMID: 23303788
- PMCID: PMC3575822
- DOI: 10.1093/nar/gks1454
Oxygen-dependent expression of cytochrome c oxidase subunit 4-2 gene expression is mediated by transcription factors RBPJ, CXXC5 and CHCHD2
Abstract
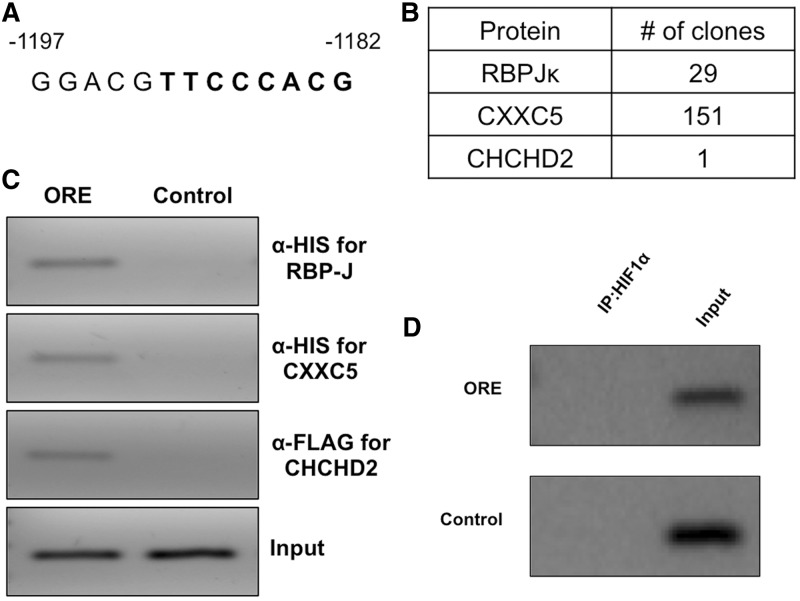
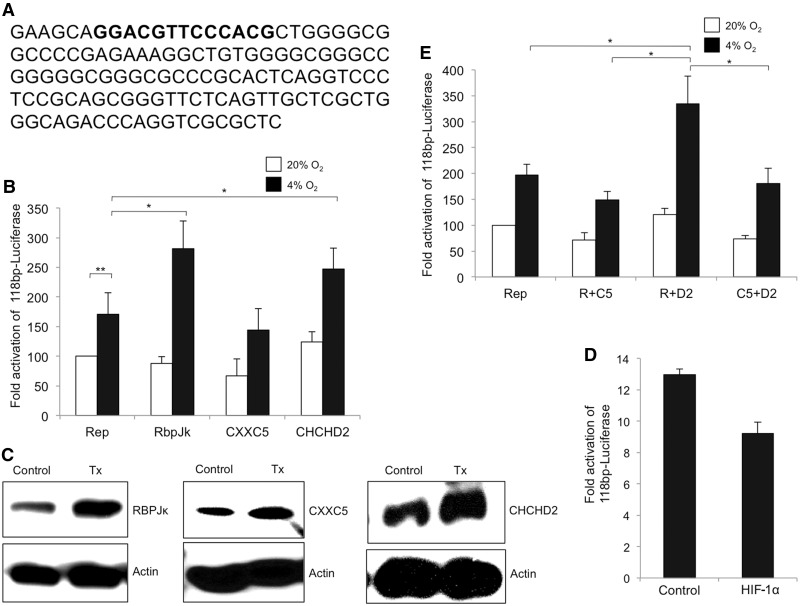
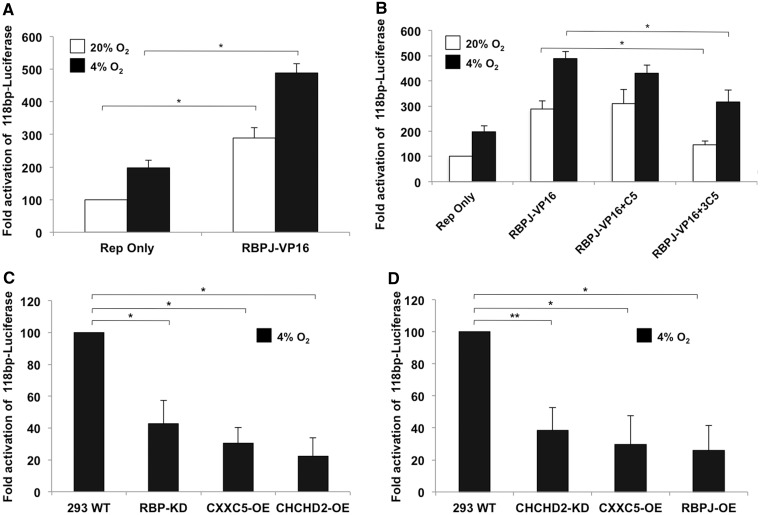
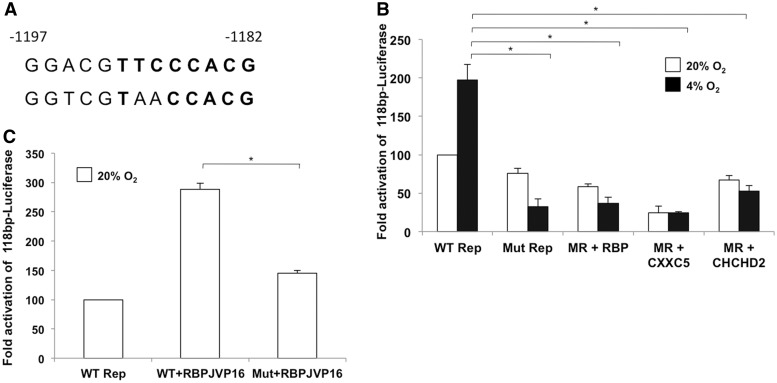
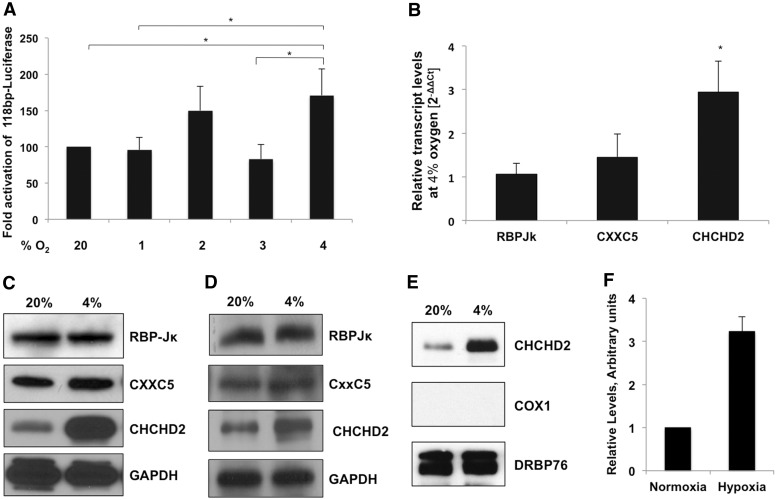
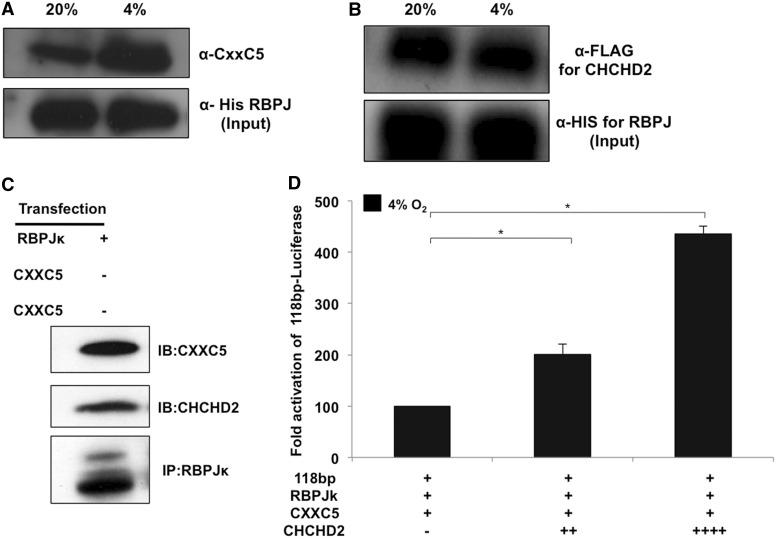
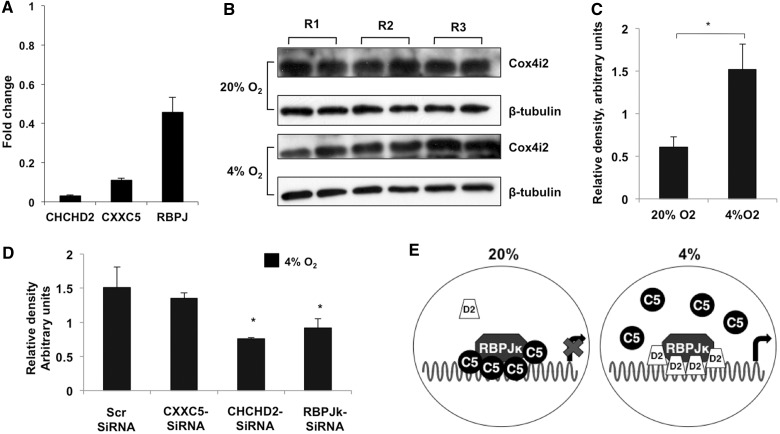
Cytochrome c oxidase (COX) is the terminal enzyme of the electron transport chain, made up of 13 subunits encoded by both mitochondrial and nuclear DNA. Subunit 4 (COX4), a key regulatory subunit, exists as two isoforms, the ubiquitous isoform 1 and the tissue-specific (predominantly lung) isoform 2 (COX4I2). COX4I2 renders lung COX about 2-fold more active compared with liver COX, which lacks COX4I2. We previously identified a highly conserved 13-bp sequence in the proximal promoter of COX4I2 that functions as an oxygen responsive element (ORE), maximally active at a 4% oxygen concentration. Here, we have identified three transcription factors that bind this conserved ORE, namely recombination signal sequence-binding protein Jκ (RBPJ), coiled-coil-helix-coiled-coil-helix domain 2 (CHCHD2) and CXXC finger protein 5 (CXXC5). We demonstrate that RBPJ and CHCHD2 function towards activating the ORE at 4% oxygen, whereas CXXC5 functions as an inhibitor. To validate results derived from cultured cells, we show using RNA interference a similar effect of these transcription factors in the gene regulation of COX4I2 in primary pulmonary arterial smooth muscle cells. Depending on the oxygen tension, a concerted action of the three transcription factors regulates the expression of COX4I2 that, as we discuss, could augment both COX activity and its ability to cope with altered cellular energy requirements.
Figures
References
-
- Napiwotzki J, Kadenbach B. Extramitochondrial ATP/ADP-ratios regulate cytochrome c oxidase activity via binding to the cytosolic domain of subunit IV. Biol. Chem. 1998;379:335–339. - PubMed
-
- Hüttemann M, Kadenbach B, Grossman LI. Mammalian subunit IV isoforms of cytochrome c oxidase. Gene. 2001;267:111–123. - PubMed
-
- Cooperstein SJ, Lazarow A. A microspectrophotometric method for the determination of cytochrome oxidase. J. Biol. Chem. 1951;189:665–670. - PubMed
-
- Huttemann M, Lee I, Liu J, Grossman LI. Transcription of mammalian cytochrome c oxidase subunit IV-2 is controlled by a novel conserved oxygen responsive element. FEBS J. 2007;274:5737–5748. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
