

Retrograde bone morphogenetic protein signaling shapes a key circadian pacemaker circuit
- PMID: 23303947
- PMCID: PMC3711639
- DOI: 10.1523/JNEUROSCI.3448-12.2013
Retrograde bone morphogenetic protein signaling shapes a key circadian pacemaker circuit
Abstract
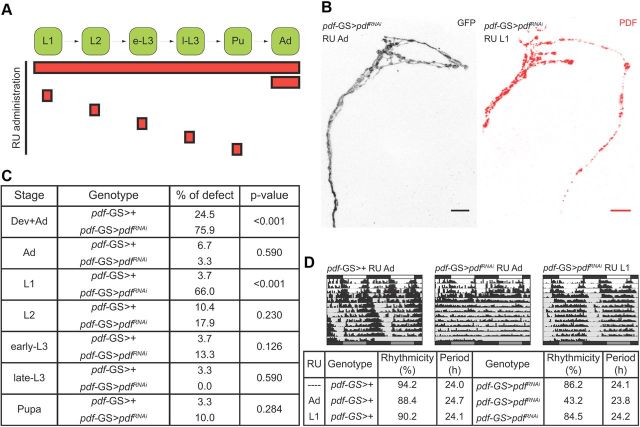
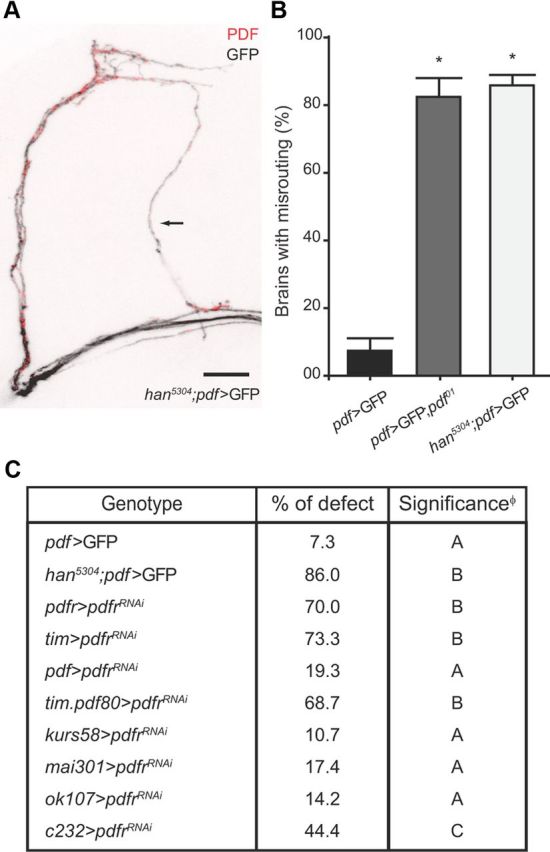
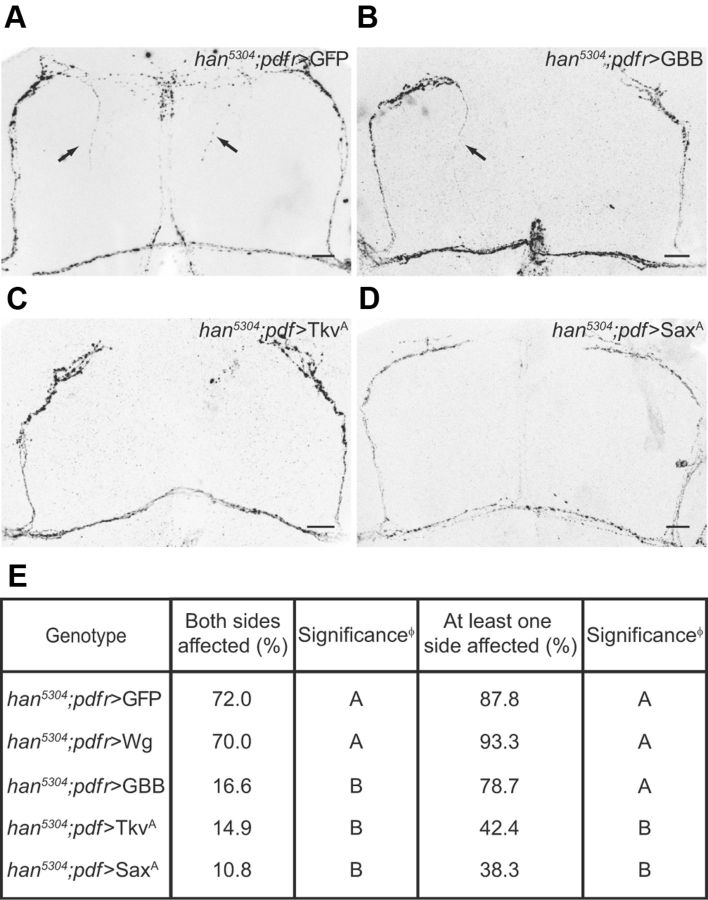
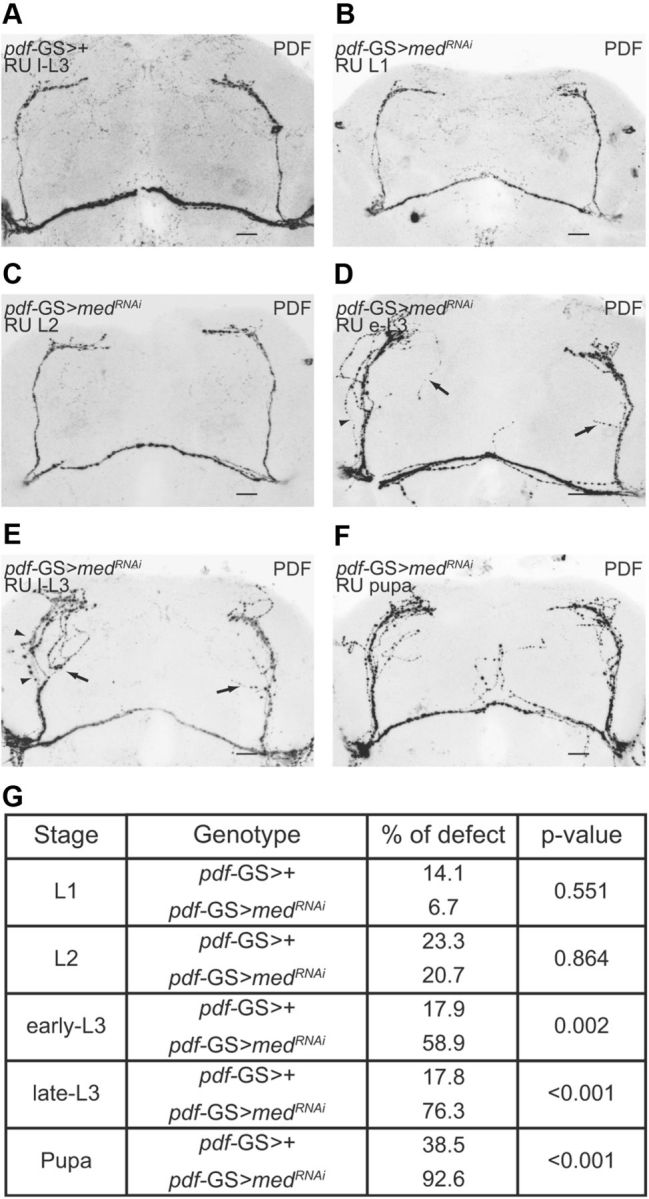
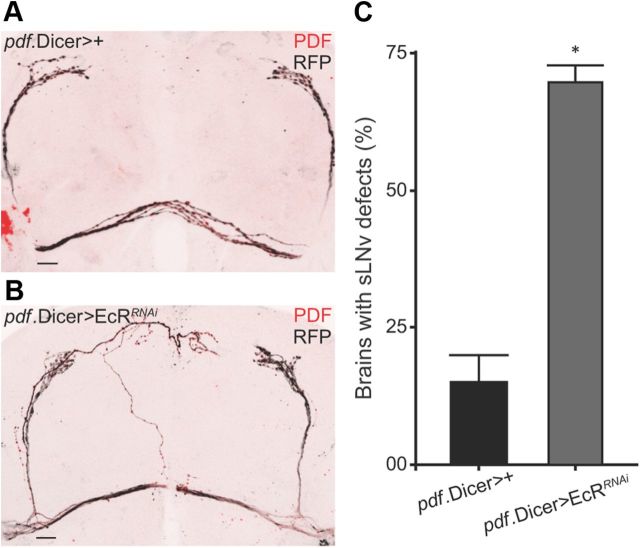
The neuropeptide pigment-dispersing factor (PDF) synchronizes molecular oscillations within circadian pacemakers in the Drosophila brain. It is expressed in the small ventral lateral neurons (sLNvs) and large ventral lateral neurons, the former being indispensable for maintaining behavioral rhythmicity under free-running conditions. How PDF circuits develop the specific connectivity traits that endow such global behavioral control remains unknown. Here, we show that mature sLNv circuits require PDF signaling during early development, acting through its cognate receptor PDFR at postsynaptic targets. Yet, axonal defects by PDF knockdown are presynaptic and become apparent only after metamorphosis, highlighting a delayed response to a signal released early on. Presynaptic expression of constitutively active bone morphogenetic protein (BMP) receptors prevents pdfr mutants misrouting phenotype, while sLNv-restricted downregulation of BMP signaling components phenocopied pdf(01). Thus, we have uncovered a novel mechanism that provides an early "tagging" of synaptic targets that will guide circuit refinement later in development.
Figures

Similar articles
-
E and M circadian pacemaker neurons use different PDF receptor signalosome components in drosophila.J Biol Rhythms. 2013 Aug;28(4):239-48. doi: 10.1177/0748730413497179. J Biol Rhythms. 2013. PMID: 23929551 Free PMC article.
-
The circadian neuropeptide PDF signals preferentially through a specific adenylate cyclase isoform AC3 in M pacemakers of Drosophila.PLoS Biol. 2012;10(6):e1001337. doi: 10.1371/journal.pbio.1001337. Epub 2012 Jun 5. PLoS Biol. 2012. PMID: 22679392 Free PMC article.
-
PDF receptor signaling in Drosophila contributes to both circadian and geotactic behaviors.Neuron. 2005 Oct 20;48(2):213-9. doi: 10.1016/j.neuron.2005.09.009. Neuron. 2005. PMID: 16242402
-
Mechanisms of clock output in the Drosophila circadian pacemaker system.J Biol Rhythms. 2006 Dec;21(6):445-57. doi: 10.1177/0748730406293910. J Biol Rhythms. 2006. PMID: 17107935 Review.
-
How does the circadian clock send timing information to the brain?Semin Cell Dev Biol. 2001 Aug;12(4):329-41. doi: 10.1006/scdb.2001.0260. Semin Cell Dev Biol. 2001. PMID: 11463217 Review.
Cited by
-
Evaluating the Autonomy of the Drosophila Circadian Clock in Dissociated Neuronal Culture.Front Cell Neurosci. 2017 Oct 12;11:317. doi: 10.3389/fncel.2017.00317. eCollection 2017. Front Cell Neurosci. 2017. PMID: 29075180 Free PMC article.
-
A genetic RNAi screen for IP₃/Ca²⁺ coupled GPCRs in Drosophila identifies the PdfR as a regulator of insect flight.PLoS Genet. 2013;9(10):e1003849. doi: 10.1371/journal.pgen.1003849. Epub 2013 Oct 3. PLoS Genet. 2013. PMID: 24098151 Free PMC article.
-
The logic of circadian organization in Drosophila.Curr Biol. 2014 Oct 6;24(19):2257-66. doi: 10.1016/j.cub.2014.08.023. Epub 2014 Sep 11. Curr Biol. 2014. PMID: 25220056 Free PMC article.
-
Mmp1 processing of the PDF neuropeptide regulates circadian structural plasticity of pacemaker neurons.PLoS Genet. 2014 Oct 30;10(10):e1004700. doi: 10.1371/journal.pgen.1004700. eCollection 2014 Oct. PLoS Genet. 2014. PMID: 25356918 Free PMC article.
-
Circadian rhythms are more resilient to pacemaker neuron disruption in female Drosophila.PLoS Biol. 2025 May 6;23(5):e3003146. doi: 10.1371/journal.pbio.3003146. eCollection 2025 May. PLoS Biol. 2025. PMID: 40327674 Free PMC article.
References
-
- Aberle H, Haghighi AP, Fetter RD, McCabe BD, Magalhães TR, Goodman CS. wishful thinking encodes a BMP type II receptor that regulates synaptic growth in Drosophila. Neuron. 2002;33:545–558. - PubMed
-
- Atwood HL, Govind CK, Wu CF. Differential ultrastructure of synaptic terminals on ventral longitudinal abdominal muscles in Drosophila larvae. J Neurobiol. 1993;24:1008–1024. - PubMed
-
- Awasaki T, Ito K. Engulfing action of glial cells is required for programmed axon pruning during Drosophila metamorphosis. Curr Biol. 2004;14:668–677. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases