

Responses of vestibular nucleus neurons to inputs from the hindlimb are enhanced following a bilateral labyrinthectomy
- PMID: 23305979
- PMCID: PMC3615601
- DOI: 10.1152/japplphysiol.01389.2012
Responses of vestibular nucleus neurons to inputs from the hindlimb are enhanced following a bilateral labyrinthectomy
Abstract
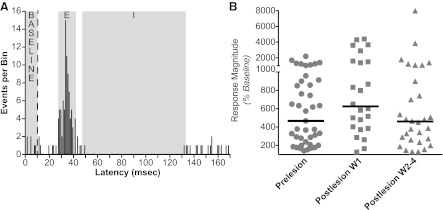
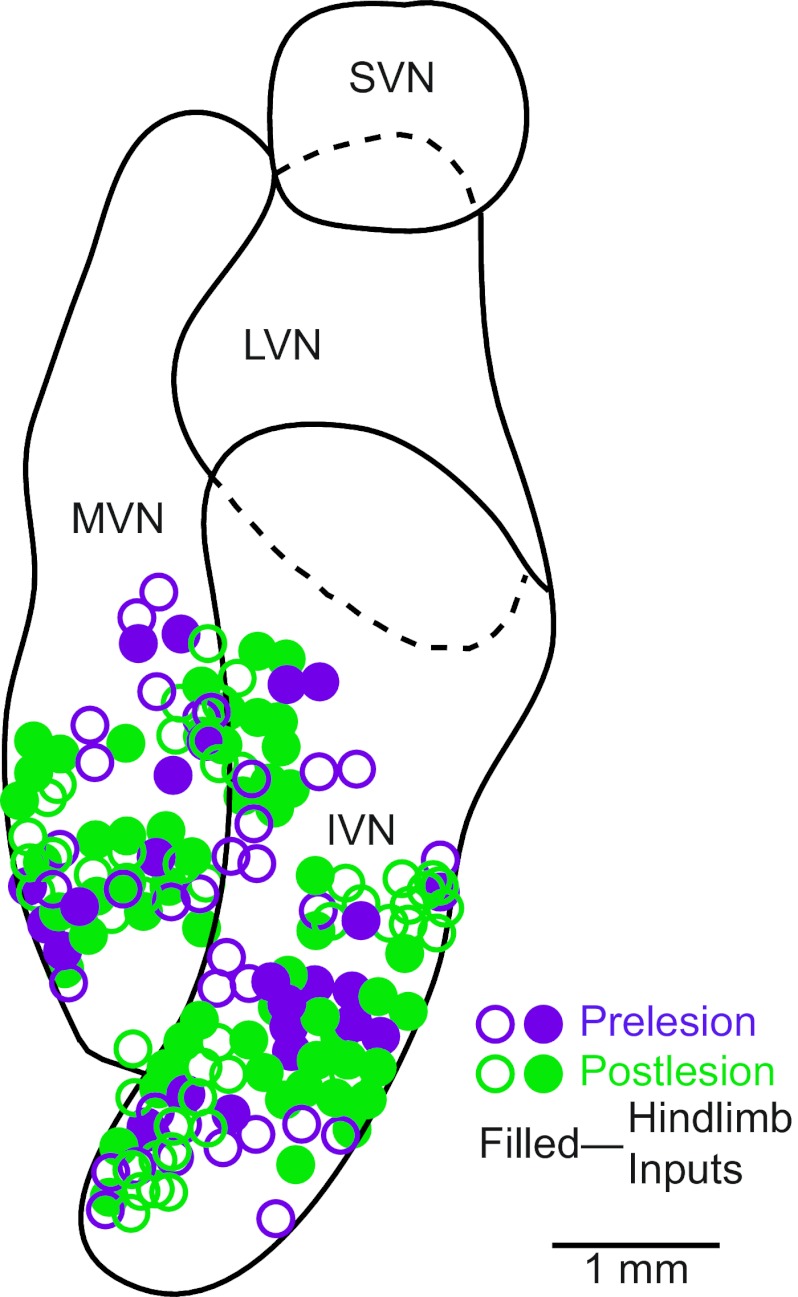
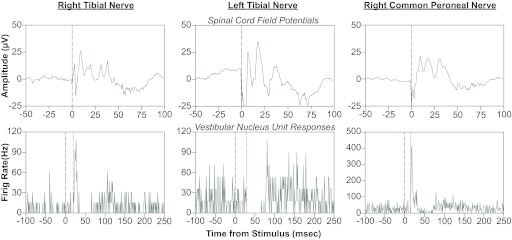
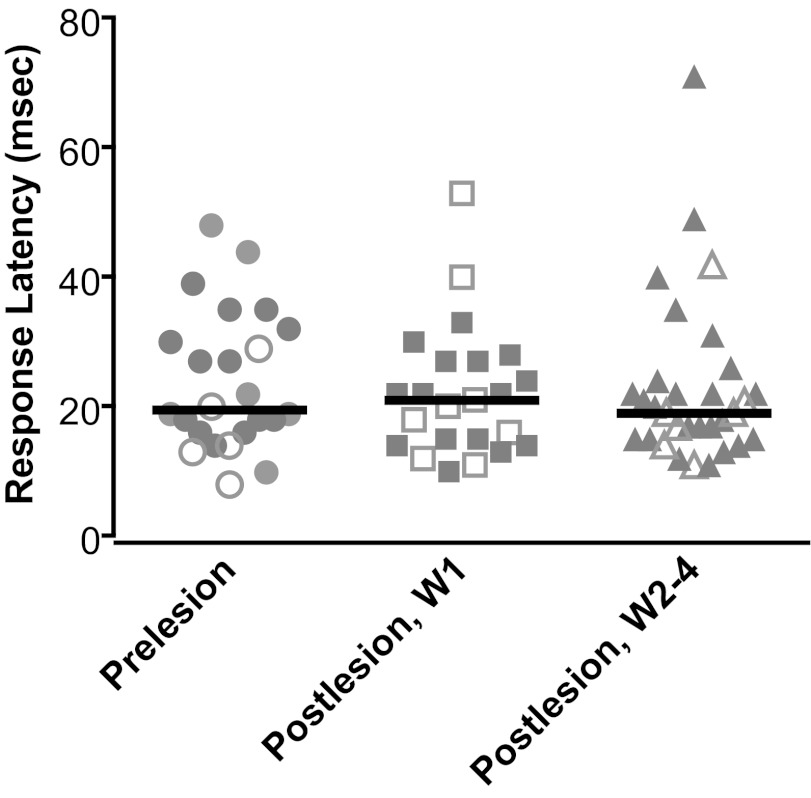
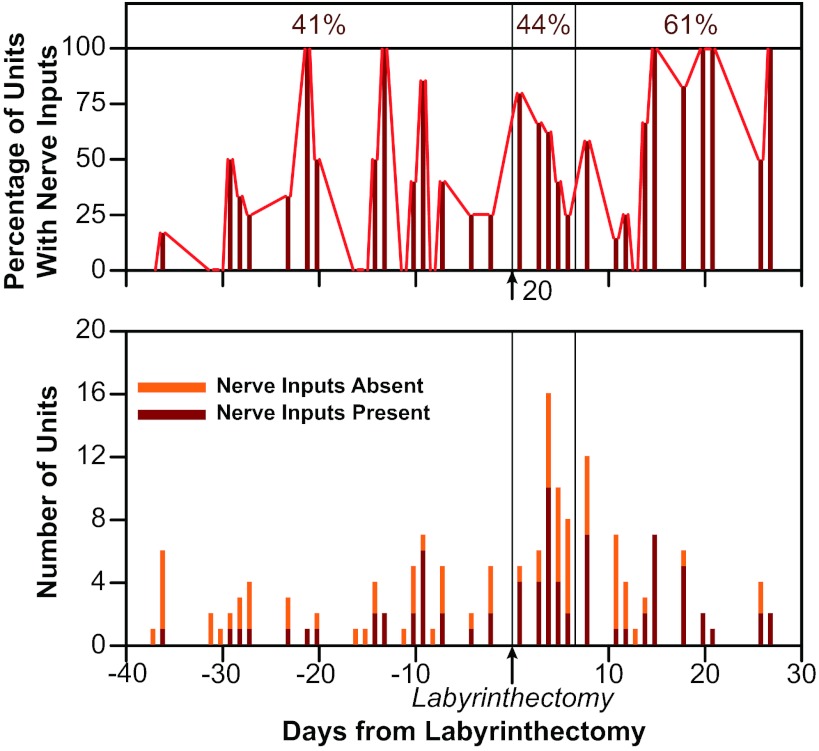
Vestibular nucleus neurons have been shown to respond to stimulation of afferents innervating the limbs. However, a limitation in the potential translation of these findings is that they were obtained from decerebrate or anesthetized animals. The goal of the present study was to determine whether stimulation of hindlimb nerves similarly affects vestibular nucleus neuronal activity in conscious cats, and whether the responsiveness of neurons to the stimuli is altered following a bilateral labyrinthectomy. In labyrinth-intact animals, the firing rate of 24/59 (41%) of the neurons in the caudal vestibular nucleus complex was affected by hindlimb nerve stimulation. Most responses were excitatory; the median response latency was 20 ms, but some units had response latencies as short as 10 ms. In the first week after a bilateral labyrinthectomy, the proportion of vestibular nucleus neurons that responded to hindlimb nerve stimulation increased slightly (to 24/55 or 44% of units). However, during the subsequent postlabyrinthectomy survival period, the proportion of vestibular nucleus neurons with hindlimb inputs increased significantly (to 30/49 or 61% of units). Stimuli to hindlimb nerves needed to elicit neuronal responses was consistently over three times the threshold for eliciting an afferent volley. These data show that inputs from hindlimb afferents smaller than those innervating muscle spindles and Golgi tendon organs affect the processing of information in the vestibular nuclei, and that these inputs are enhanced following a bilateral labyrinthectomy. These findings have implications for the development of a limb neuroprosthetics device for the management of bilateral vestibular loss.
Figures
References
-
- Anastasopoulos D, Mergner T. Canal-neck interaction in vestibular nuclear neurons of the cat. Exp Brain Res 46: 269–280, 1982 - PubMed
-
- Andersson G, Oscarsson O. Climbing fiber microzones in cerebellar vermis and their projection to different groups of cells in the lateral vestibular nucleus. Exp Brain Res 32: 565–579, 1978 - PubMed
-
- Baker J, Goldberg J, Hermann G, Peterson B. Spatial and temporal response properties of secondary neurons that receive convergent input in vestibular nuclei of alert cats. Brain Res 294: 138–143, 1984 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous