

Tissue-specific control of brain-enriched miR-7 biogenesis
- PMID: 23307866
- PMCID: PMC3553281
- DOI: 10.1101/gad.199190.112
Tissue-specific control of brain-enriched miR-7 biogenesis
Abstract
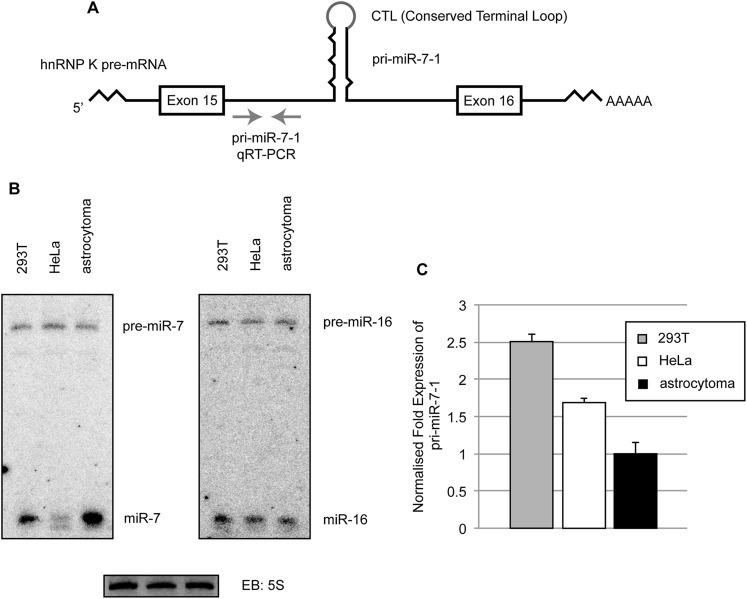
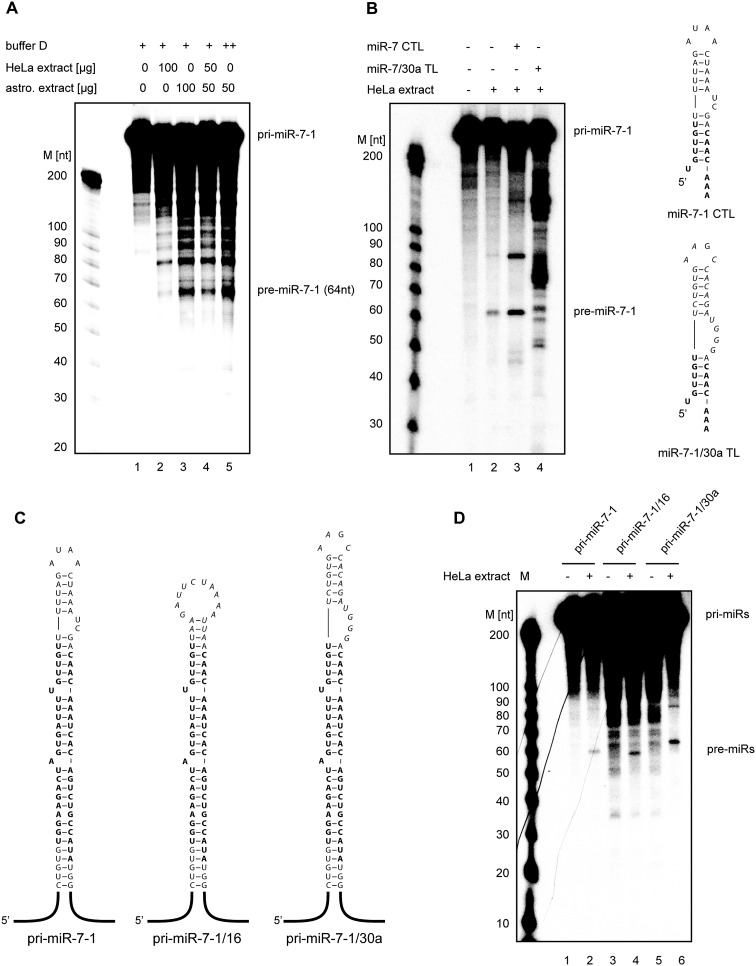
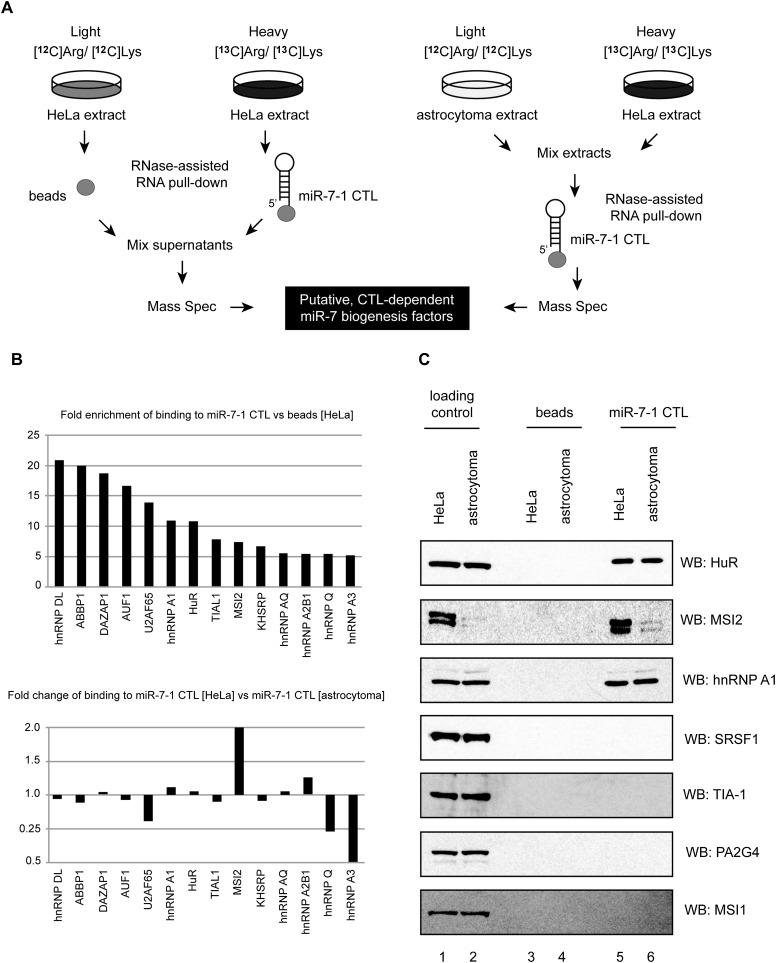
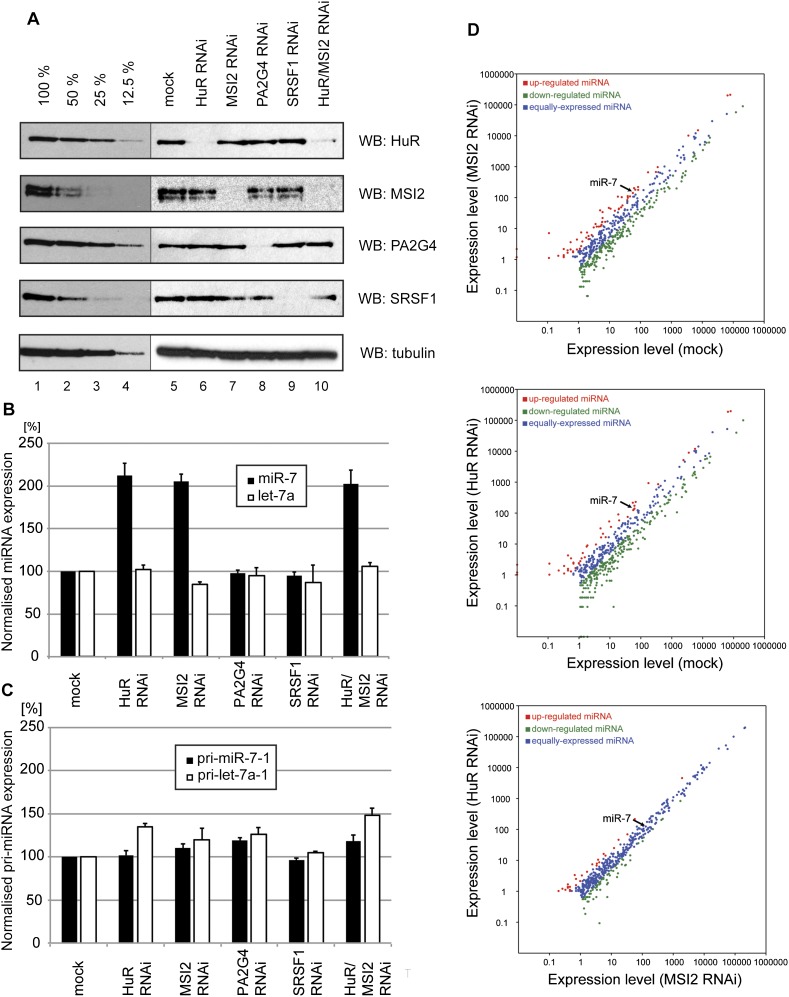
MicroRNA (miRNA) biogenesis is a highly regulated process in eukaryotic cells. Several mature miRNAs exhibit a tissue-specific pattern of expression without an apparent tissue-specific pattern for their corresponding primary transcripts. This discrepancy is suggestive of post-transcriptional regulation of miRNA abundance. Here, we demonstrate that the brain-enriched expression of miR-7, which is processed from the ubiquitous hnRNP K pre-mRNA transcript, is achieved by inhibition of its biogenesis in nonbrain cells in both human and mouse systems. Using stable isotope labeling by amino acids in cell culture (SILAC) mass spectrometry combined with RNase-assisted RNA pull-down, we identified Musashi homolog 2 (MSI2) and Hu antigen R (HuR) proteins as inhibitors of miR-7 processing in nonneural cells. This is achieved through HuR-mediated binding of MSI2 to the conserved terminal loop of pri-miR-7. Footprinting and electrophoretic gel mobility shift analysis (EMSA) provide further evidence for a direct interaction between pri-miR-7-1 and the HuR/MSI2 complex, resulting in stabilization of the pri-miR-7-1 structure. We also confirmed the physiological relevance of this inhibitory mechanism in a neuronal differentiation system using human SH-SY5Y cells. Finally, we show elevated levels of miR-7 in selected tissues from MSI2 knockout (KO) mice without apparent changes in the abundance of the pri-miR-7 transcript. Altogether, our data provide the first insight into the regulation of brain-enriched miRNA processing by defined tissue-specific factors.
Figures
Similar articles
-
Oleic Acid Induces MiR-7 Processing through Remodeling of Pri-MiR-7/Protein Complex.J Mol Biol. 2017 Jun 2;429(11):1638-1649. doi: 10.1016/j.jmb.2017.05.001. Epub 2017 May 5. J Mol Biol. 2017. PMID: 28483648 Free PMC article.
-
Msi2-mediated MiR7a-1 processing repression promotes myogenesis.J Cachexia Sarcopenia Muscle. 2022 Feb;13(1):728-742. doi: 10.1002/jcsm.12882. Epub 2021 Dec 8. J Cachexia Sarcopenia Muscle. 2022. PMID: 34877814 Free PMC article.
-
Polypyrimidine tract-binding protein blocks miRNA-124 biogenesis to enforce its neuronal-specific expression in the mouse.Proc Natl Acad Sci U S A. 2018 Nov 20;115(47):E11061-E11070. doi: 10.1073/pnas.1809609115. Epub 2018 Nov 6. Proc Natl Acad Sci U S A. 2018. PMID: 30401736 Free PMC article.
-
Regulation of the MIR155 host gene in physiological and pathological processes.Gene. 2013 Dec 10;532(1):1-12. doi: 10.1016/j.gene.2012.12.009. Epub 2012 Dec 14. Gene. 2013. PMID: 23246696 Review.
-
Stimulation of pri-miR-18a processing by hnRNP A1.Adv Exp Med Biol. 2010;700:28-35. doi: 10.1007/978-1-4419-7823-3_3. Adv Exp Med Biol. 2010. PMID: 21627027 Review.
Cited by
-
Long non-coding RNA ELDR enhances oral cancer growth by promoting ILF3-cyclin E1 signaling.EMBO Rep. 2020 Dec 3;21(12):e51042. doi: 10.15252/embr.202051042. Epub 2020 Oct 12. EMBO Rep. 2020. PMID: 33043604 Free PMC article.
-
RNA-Targeted Therapies and High-Throughput Screening Methods.Int J Mol Sci. 2020 Apr 23;21(8):2996. doi: 10.3390/ijms21082996. Int J Mol Sci. 2020. PMID: 32340368 Free PMC article. Review.
-
RNA-Binding Protein MSI2 Binds to miR-301a-3p and Facilitates Its Distribution in Mitochondria of Endothelial Cells.Front Mol Biosci. 2021 Jan 21;7:609828. doi: 10.3389/fmolb.2020.609828. eCollection 2020. Front Mol Biosci. 2021. PMID: 33553241 Free PMC article.
-
RNA-binding activity of TRIM25 is mediated by its PRY/SPRY domain and is required for ubiquitination.BMC Biol. 2017 Nov 8;15(1):105. doi: 10.1186/s12915-017-0444-9. BMC Biol. 2017. PMID: 29117863 Free PMC article.
-
Quantitative identification of proteins that influence miRNA biogenesis by RNA pull-down-SILAC mass spectrometry (RP-SMS).Methods. 2019 Jan 1;152:12-17. doi: 10.1016/j.ymeth.2018.06.006. Epub 2018 Jun 8. Methods. 2019. PMID: 29890283 Free PMC article.
References
-
- Barbouti A, Hoglund M, Johansson B, Lassen C, Nilsson PG, Hagemeijer A, Mitelman F, Fioretos T 2003. A novel gene, MSI2, encoding a putative RNA-binding protein is recurrently rearranged at disease progression of chronic myeloid leukemia and forms a fusion gene with HOXA9 as a result of the cryptic t(7;17)(p15;q23). Cancer Res 63: 1202–1206 - PubMed
-
- Bushati N, Cohen SM 2007. MicroRNA functions. Annu Rev Cell Dev Biol 23: 175–205 - PubMed
-
- de Andres-Aguayo L, Varas F, Kallin EM, Infante JF, Wurst W, Floss T, Graf T 2011. Musashi 2 is a regulator of the HSC compartment identified by a retroviral insertion screen and knockout mice. Blood 118: 554–564 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous